

The Legionella pneumophila effector protein, LegC7, alters yeast endosomal trafficking
- PMID: 25643265
- PMCID: PMC4314205
- DOI: 10.1371/journal.pone.0116824
The Legionella pneumophila effector protein, LegC7, alters yeast endosomal trafficking
Abstract
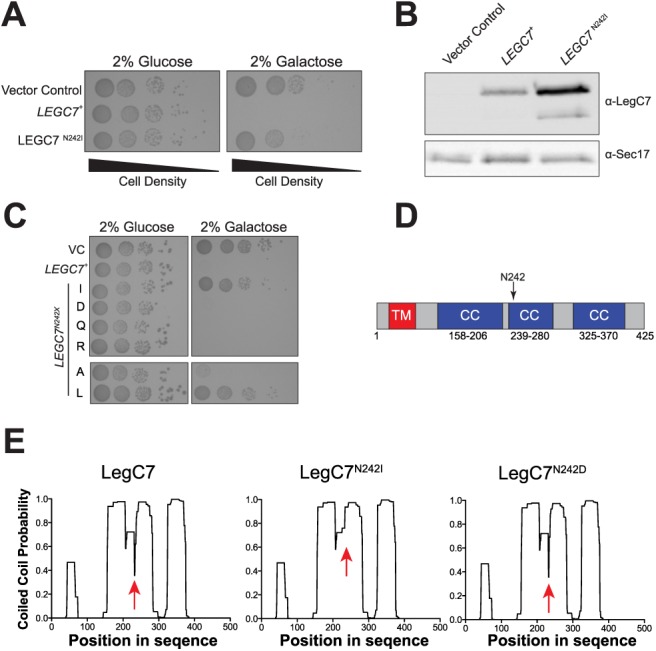
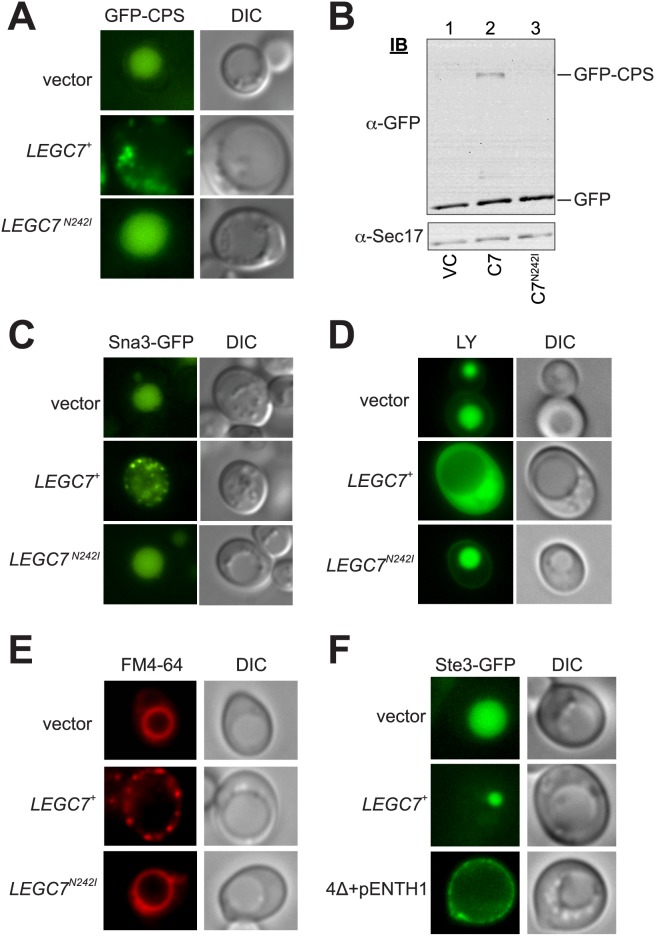
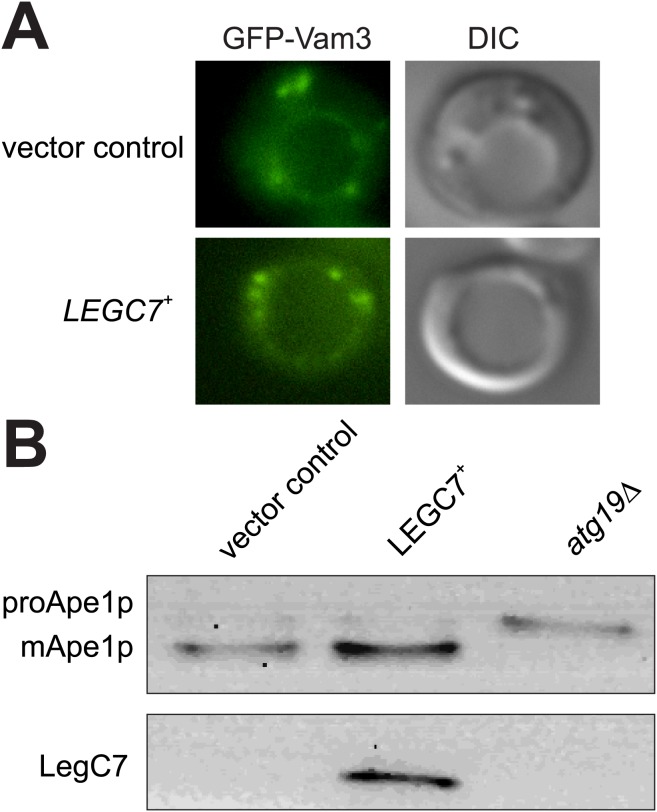
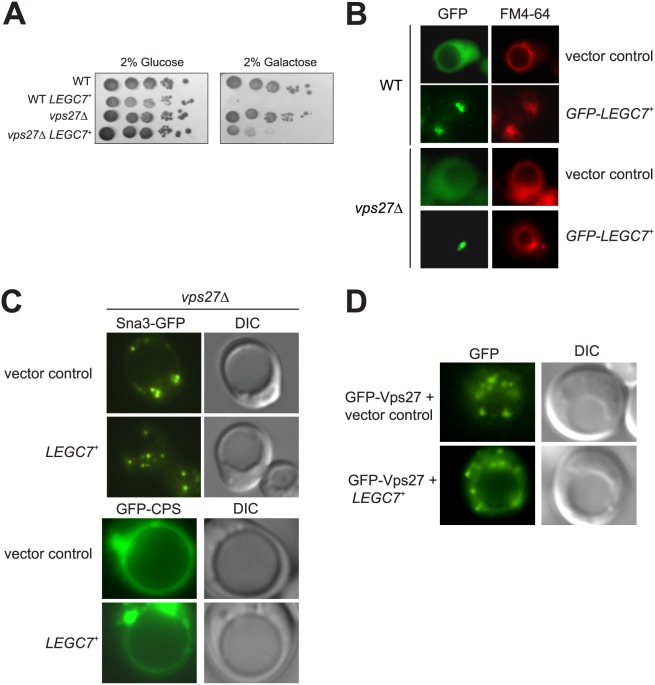
The intracellular pathogen, Legionella pneumophila, relies on numerous secreted effector proteins to manipulate host endomembrane trafficking events during pathogenesis, thereby preventing fusion of the bacteria-laden phagosome with host endolysosomal compartments, and thus escaping degradation. Upon expression in the surrogate eukaryotic model Saccharomyces cerevisiae, we find that the L. pneumophila LegC7/YlfA effector protein disrupts the delivery of both biosynthetic and endocytic cargo to the yeast vacuole. We demonstrate that the effects of LegC7 are specific to the endosome:vacuole delivery pathways; LegC7 expression does not disrupt other known vacuole-directed pathways. Deletions of the ESCRT-0 complex member, VPS27, provide resistance to the LegC7 toxicity, providing a possible target for LegC7 function in vivo. Furthermore, a single amino acid substitution in LegC7 abrogates both its toxicity and ability to alter endosomal traffic in vivo, thereby identifying a critical functional domain. LegC7 likely inhibits endosomal trafficking during L. pneumophila pathogenesis to prevent entry of the phagosome into the endosomal maturation pathway and eventual fusion with the lysosome.
Conflict of interest statement
Figures
Similar articles
-
Legionella pneumophila LegC7 effector protein drives aberrant endoplasmic reticulum:endosome contacts in yeast.Traffic. 2021 Aug;22(8):284-302. doi: 10.1111/tra.12807. Epub 2021 Jul 7. Traffic. 2021. PMID: 34184807 Free PMC article.
-
Legionella eukaryotic-like type IV substrates interfere with organelle trafficking.PLoS Pathog. 2008 Aug 1;4(8):e1000117. doi: 10.1371/journal.ppat.1000117. PLoS Pathog. 2008. PMID: 18670632 Free PMC article.
-
LegC3, an effector protein from Legionella pneumophila, inhibits homotypic yeast vacuole fusion in vivo and in vitro.PLoS One. 2013;8(2):e56798. doi: 10.1371/journal.pone.0056798. Epub 2013 Feb 20. PLoS One. 2013. PMID: 23437241 Free PMC article.
-
Formation of the Legionella Replicative Compartment at the Crossroads of Retrograde Trafficking.Front Cell Infect Microbiol. 2017 Nov 24;7:482. doi: 10.3389/fcimb.2017.00482. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 29226112 Free PMC article. Review.
-
Tethering complexes in the endocytic pathway: CORVET and HOPS.FEBS J. 2013 Jun;280(12):2743-57. doi: 10.1111/febs.12151. Epub 2013 Feb 21. FEBS J. 2013. PMID: 23351085 Review.
Cited by
-
The Legionella pneumophila genome evolved to accommodate multiple regulatory mechanisms controlled by the CsrA-system.PLoS Genet. 2017 Feb 17;13(2):e1006629. doi: 10.1371/journal.pgen.1006629. eCollection 2017 Feb. PLoS Genet. 2017. PMID: 28212376 Free PMC article.
-
The Toxoplasma Vacuolar H+-ATPase Regulates Intracellular pH and Impacts the Maturation of Essential Secretory Proteins.Cell Rep. 2019 May 14;27(7):2132-2146.e7. doi: 10.1016/j.celrep.2019.04.038. Cell Rep. 2019. PMID: 31091451 Free PMC article.
-
An Indispensable Role for the MavE Effector of Legionella pneumophila in Lysosomal Evasion.mBio. 2021 Feb 9;12(1):e03458-20. doi: 10.1128/mBio.03458-20. mBio. 2021. PMID: 33563829 Free PMC article.
-
Molecular Mimicry: a Paradigm of Host-Microbe Coevolution Illustrated by Legionella.mBio. 2020 Oct 6;11(5):e01201-20. doi: 10.1128/mBio.01201-20. mBio. 2020. PMID: 33024033 Free PMC article. Review.
-
Idiosyncratic Biogenesis of Intracellular Pathogens-Containing Vacuoles.Front Cell Infect Microbiol. 2021 Nov 11;11:722433. doi: 10.3389/fcimb.2021.722433. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34858868 Free PMC article. Review.
References
-
- Kaufmann AF, Mcdade JE, Patton CE, Bennett JV, Skaliy P, et al. (1981) Pontiac Fever: Isolation Of The Etiologic Agent (Legionella Pneumophila) And Demonstration Of Its Mode Of Transmission. American Journal of Epidemiology 114: 337–347. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
