

Epigenetic basis of opiate suppression of Bdnf gene expression in the ventral tegmental area
- PMID: 25643298
- PMCID: PMC4340719
- DOI: 10.1038/nn.3932
Epigenetic basis of opiate suppression of Bdnf gene expression in the ventral tegmental area
Abstract
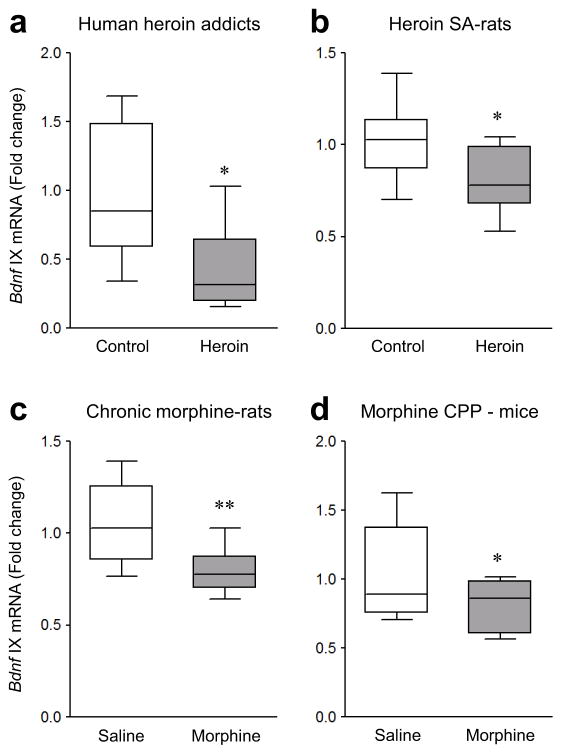
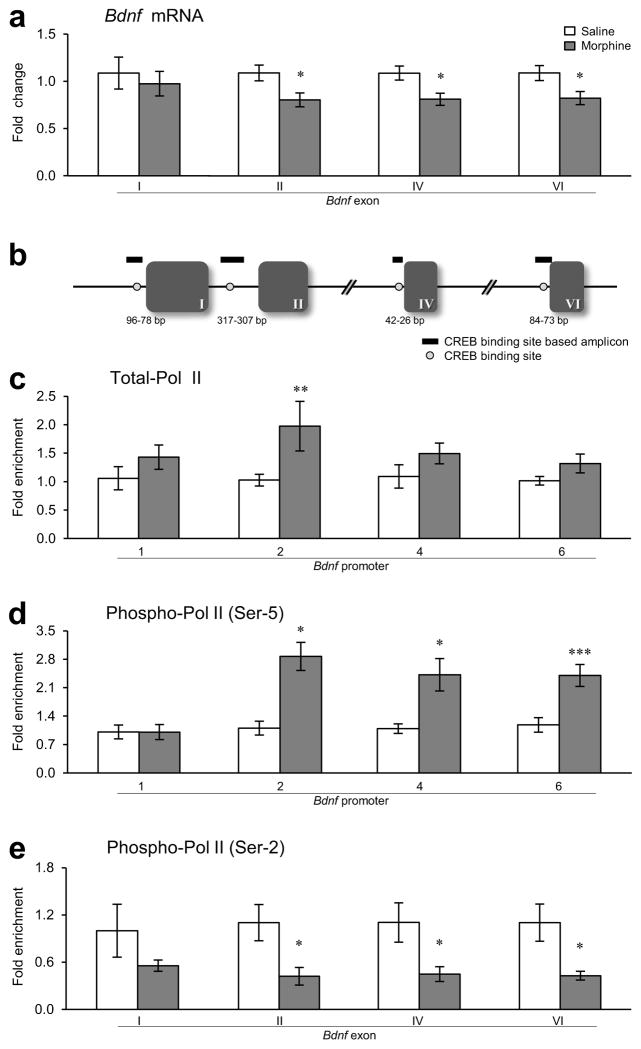
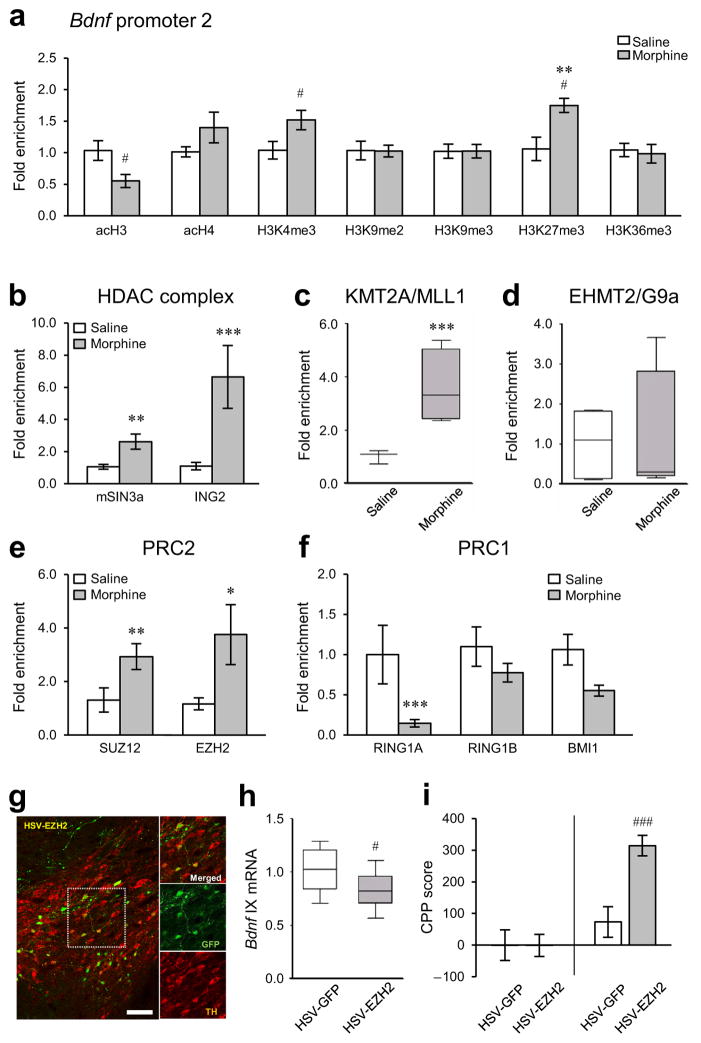
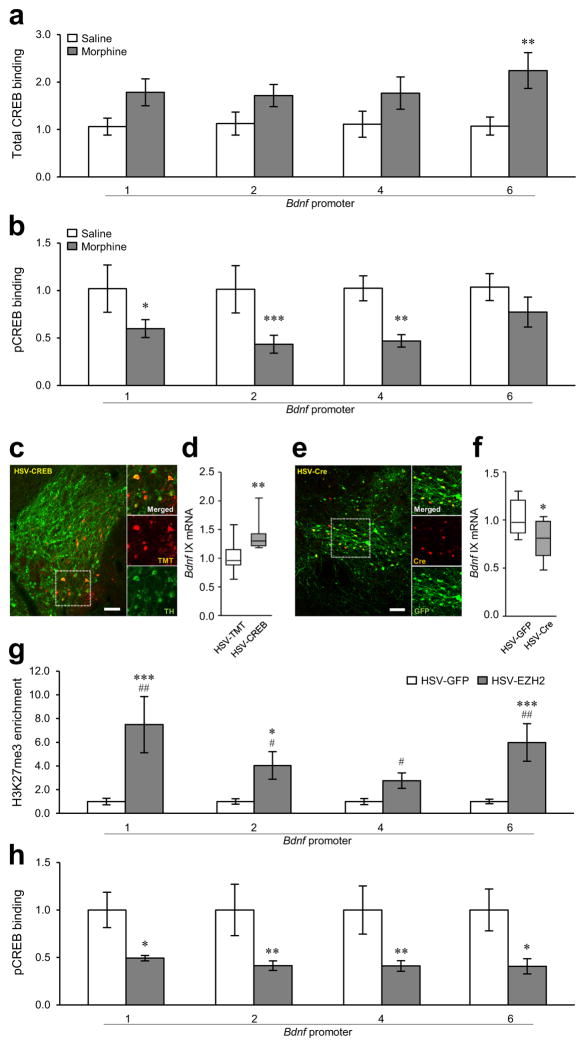
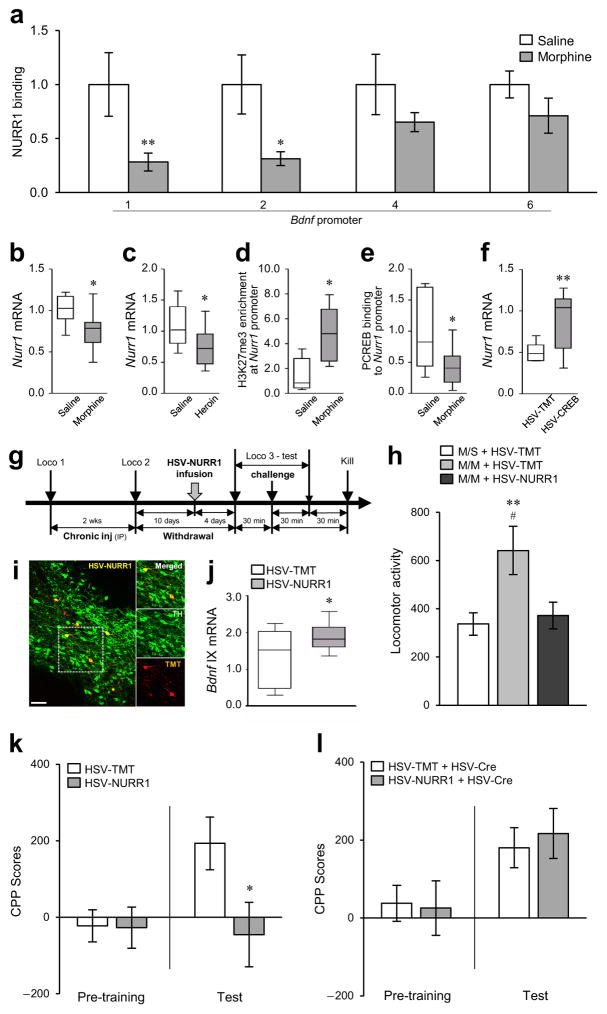
Brain-derived neurotrophic factor (BDNF) has a crucial role in modulating neural and behavioral plasticity to drugs of abuse. We found a persistent downregulation of exon-specific Bdnf expression in the ventral tegmental area (VTA) in response to chronic opiate exposure, which was mediated by specific epigenetic modifications at the corresponding Bdnf gene promoters. Exposure to chronic morphine increased stalling of RNA polymerase II at these Bdnf promoters in VTA and altered permissive and repressive histone modifications and occupancy of their regulatory proteins at the specific promoters. Furthermore, we found that morphine suppressed binding of phospho-CREB (cAMP response element binding protein) to Bdnf promoters in VTA, which resulted from enrichment of trimethylated H3K27 at the promoters, and that decreased NURR1 (nuclear receptor related-1) expression also contributed to Bdnf repression and associated behavioral plasticity to morphine. Our findings suggest previously unknown epigenetic mechanisms of morphine-induced molecular and behavioral neuroadaptations.
Conflict of interest statement
Competing Financial Interests: The authors declare no competing financial interests.
Figures
Comment in
-
Gene expression: Putting a stop to BDNF.Nat Rev Neurosci. 2015 Apr;16(4):186-7. doi: 10.1038/nrn3933. Epub 2015 Mar 4. Nat Rev Neurosci. 2015. PMID: 25736021 No abstract available.
References
-
- Pu L, Liu QS, Poo MM. BDNF-dependent synaptic sensitization in midbrain dopamine neurons after cocaine withdrawal. Nat Neurosci. 2006;9:605–607. - PubMed
-
- Filip M, et al. Alterations in BDNF and trkB mRNAs following acute or sensitizing cocaine treatments and withdrawal. Brain Res. 2006;1071:218–225. - PubMed
-
- Graham DL, et al. Dynamic BDNF activity in nucleus accumbens with cocaine use increases self-administration and relapse. Nat Neurosci. 2007;10:1029–1037. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
