

Mechanism of microhomology-mediated end-joining promoted by human DNA polymerase θ
- PMID: 25643323
- PMCID: PMC4351179
- DOI: 10.1038/nsmb.2961
Mechanism of microhomology-mediated end-joining promoted by human DNA polymerase θ
Abstract
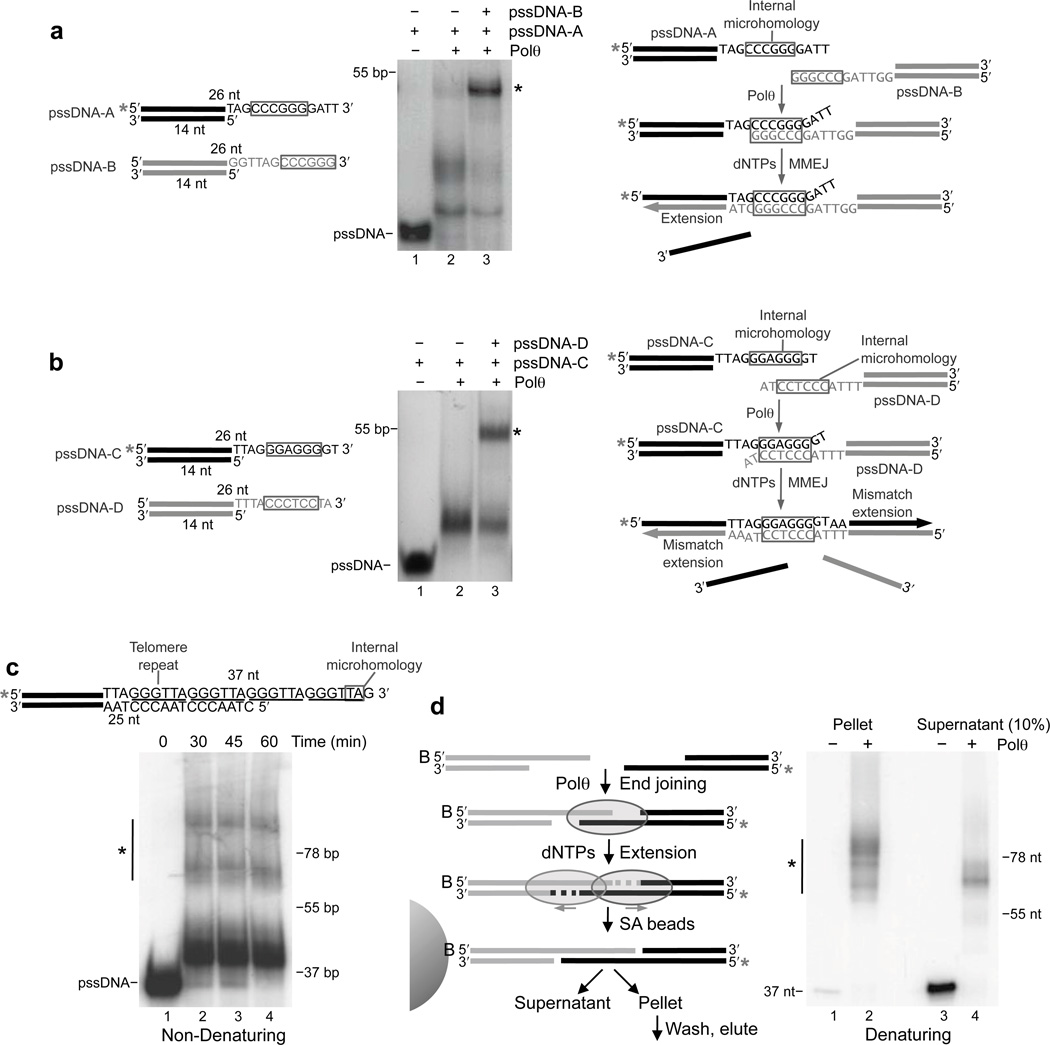
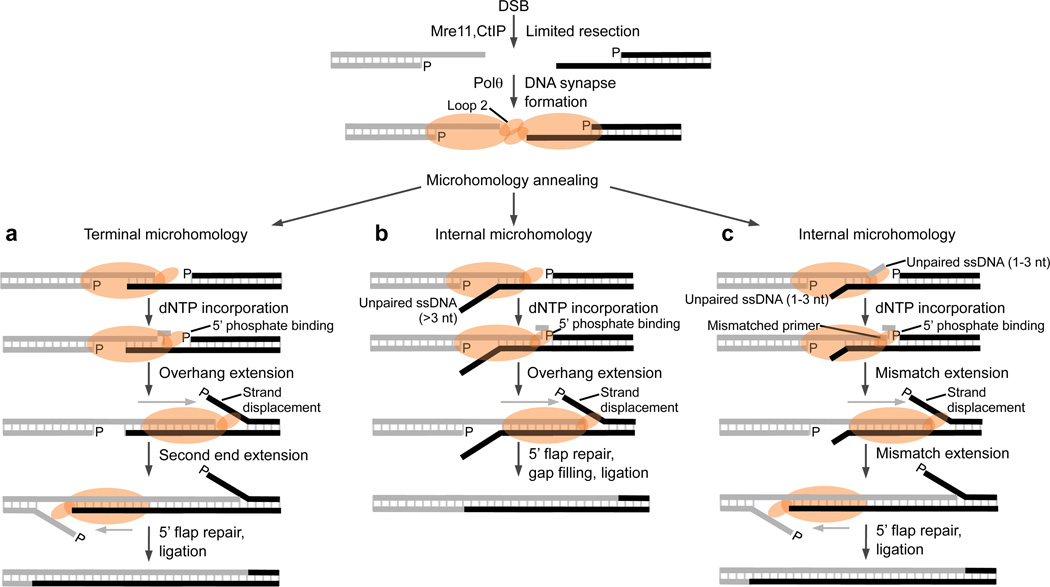
Microhomology-mediated end-joining (MMEJ) is an error-prone alternative double-strand break-repair pathway that uses sequence microhomology to recombine broken DNA. Although MMEJ has been implicated in cancer development, the mechanism of this pathway is unknown. We demonstrate that purified human DNA polymerase θ (Polθ) performs MMEJ of DNA containing 3' single-strand DNA overhangs with ≥2 bp of homology, including DNA modeled after telomeres, and show that MMEJ is dependent on Polθ in human cells. Our data support a mechanism whereby Polθ facilitates end-joining and microhomology annealing, then uses the opposing overhang as a template in trans to stabilize the DNA synapse. Polθ exhibits a preference for DNA containing a 5'-terminal phosphate, similarly to polymerases involved in nonhomologous end-joining. Finally, we identify a conserved loop domain that is essential for MMEJ and higher-order structures of Polθ that probably promote DNA synapse formation.
Figures





Similar articles
-
Molecular basis of microhomology-mediated end-joining by purified full-length Polθ.Nat Commun. 2019 Sep 27;10(1):4423. doi: 10.1038/s41467-019-12272-9. Nat Commun. 2019. PMID: 31562312 Free PMC article.
-
Structural basis for Polθ-helicase DNA binding and microhomology-mediated end-joining.Nat Commun. 2025 Apr 19;16(1):3725. doi: 10.1038/s41467-025-58441-x. Nat Commun. 2025. PMID: 40253368 Free PMC article.
-
Polymerase theta is a synthetic lethal target for killing Epstein-Barr virus lymphomas.J Virol. 2024 Jul 23;98(7):e0057224. doi: 10.1128/jvi.00572-24. Epub 2024 Jun 11. J Virol. 2024. PMID: 38860782 Free PMC article.
-
Microhomology-Mediated End Joining: A Back-up Survival Mechanism or Dedicated Pathway?Trends Biochem Sci. 2015 Nov;40(11):701-714. doi: 10.1016/j.tibs.2015.08.006. Epub 2015 Oct 1. Trends Biochem Sci. 2015. PMID: 26439531 Free PMC article. Review.
-
Risky business: Microhomology-mediated end joining.Mutat Res. 2016 Jun;788:17-24. doi: 10.1016/j.mrfmmm.2015.12.005. Epub 2016 Jan 2. Mutat Res. 2016. PMID: 26790771 Free PMC article. Review.
Cited by
-
Zebrafish Polymerase Theta and human Polymerase Theta: orthologues with homologous function.bioRxiv [Preprint]. 2024 Oct 7:2024.09.27.615541. doi: 10.1101/2024.09.27.615541. bioRxiv. 2024. Update in: PLoS One. 2025 Apr 29;20(4):e0321886. doi: 10.1371/journal.pone.0321886. PMID: 39386538 Free PMC article. Updated. Preprint.
-
DNA Repair Pathway Choices in CRISPR-Cas9-Mediated Genome Editing.Trends Genet. 2021 Jul;37(7):639-656. doi: 10.1016/j.tig.2021.02.008. Epub 2021 Apr 22. Trends Genet. 2021. PMID: 33896583 Free PMC article. Review.
-
Repair of DNA double-strand breaks by mammalian alternative end-joining pathways.J Biol Chem. 2018 Jul 6;293(27):10536-10546. doi: 10.1074/jbc.TM117.000375. Epub 2018 Mar 12. J Biol Chem. 2018. PMID: 29530982 Free PMC article. Review.
-
Mechanisms for Complex Chromosomal Insertions.PLoS Genet. 2016 Nov 23;12(11):e1006446. doi: 10.1371/journal.pgen.1006446. eCollection 2016 Nov. PLoS Genet. 2016. PMID: 27880765 Free PMC article.
-
POLQ suppresses genome instability and alterations in DNA repeat tract lengths.NAR Cancer. 2022 Jun 29;4(3):zcac020. doi: 10.1093/narcan/zcac020. eCollection 2022 Sep. NAR Cancer. 2022. PMID: 35774233 Free PMC article.
References
METHODS REFERENCES
-
- Gunn A, Stark JM. I-SceI-based assays to examine distinct repair outcomes of mammalian chromosomal double strand breaks. Methods Mol Biol. 2012;920:379–391. - PubMed
-
- Guex N, Peitsch MC. SWISS-MODEL and the Swiss-PdbViewer: an environment for comparative protein modeling. Electrophoresis. 1997;18:2714–2723. - PubMed
-
- DeLano WL. Unraveling hot spots in binding interfaces: progress and challenges. Curr Opin Struct Biol. 2002;12:14–20. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
