

The flagellar regulator TviA reduces pyroptosis by Salmonella enterica serovar Typhi
- PMID: 25644011
- PMCID: PMC4363433
- DOI: 10.1128/IAI.02803-14
The flagellar regulator TviA reduces pyroptosis by Salmonella enterica serovar Typhi
Abstract
To discern virulent from innocuous microbes, the innate immune system senses events associated with bacterial access to immunoprivileged sites such as the host cell cytosol. One such pathway is triggered by the cytosolic delivery of flagellin, the major subunit of the flagellum, by bacterial secretion systems. This leads to inflammasome activation and subsequent proinflammatory cell death (pyroptosis) of the infected phagocyte. In this study, we demonstrate that the causative agent of typhoid fever, Salmonella enterica serovar Typhi, can partially subvert this critical innate immune recognition event. The transcriptional regulator TviA, which is absent from Salmonella serovars associated with human gastroenteritis, repressed the expression of flagellin during infection of human macrophage-like (THP-1) cells. This mechanism allowed S. Typhi to dampen inflammasome activation, leading to reduced interleukin-1β (IL-1β) secretion and diminished cell death. Likewise, the introduction of the tviA gene in nontyphoidal Salmonella enterica serovar Typhimurium reduced flagellin-induced pyroptosis. These data suggest that gene regulation of virulence factors enables S. Typhi to evade innate immune recognition by concealing a pathogen-induced process from being sensed by the inflammasome.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
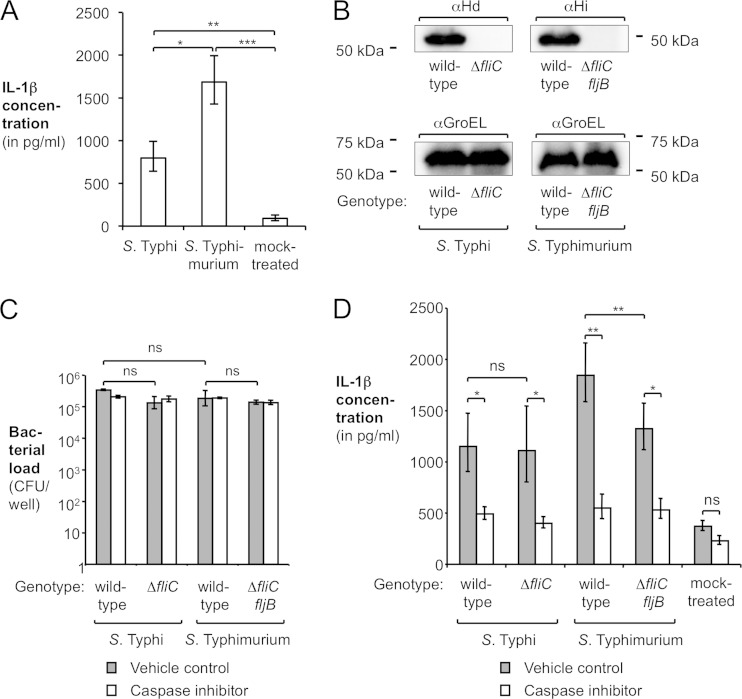
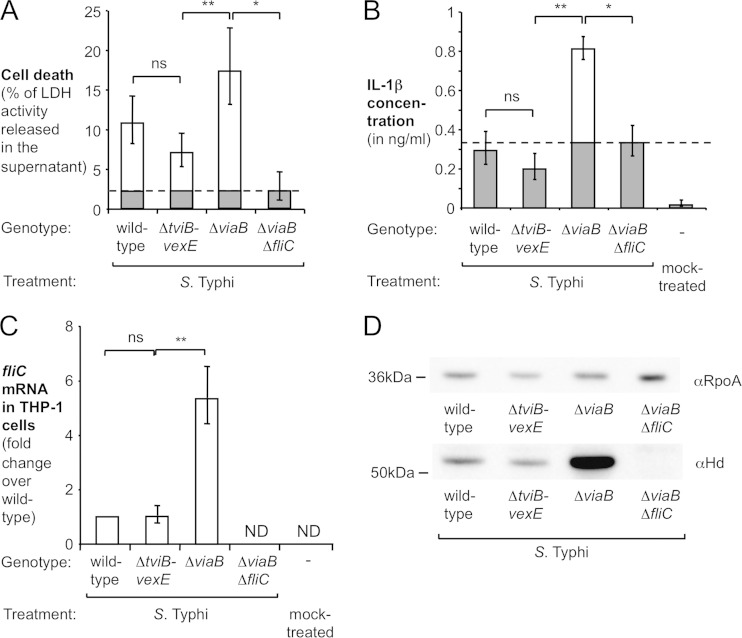
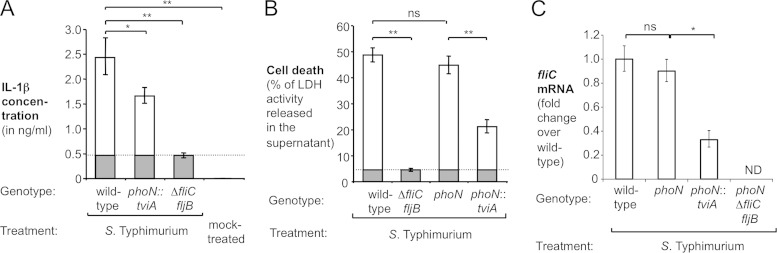
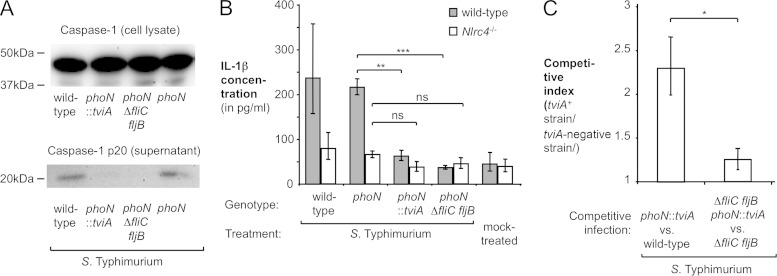
References
-
- Janeway CA., Jr 1989. Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harb Symp Quant Biol 54(Part 1):1–13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
