

Cell cycle deregulation and mosaic loss of Ext1 drive peripheral chondrosarcomagenesis in the mouse and reveal an intrinsic cilia deficiency
- PMID: 25644707
- PMCID: PMC4564123
- DOI: 10.1002/path.4510
Cell cycle deregulation and mosaic loss of Ext1 drive peripheral chondrosarcomagenesis in the mouse and reveal an intrinsic cilia deficiency
Abstract
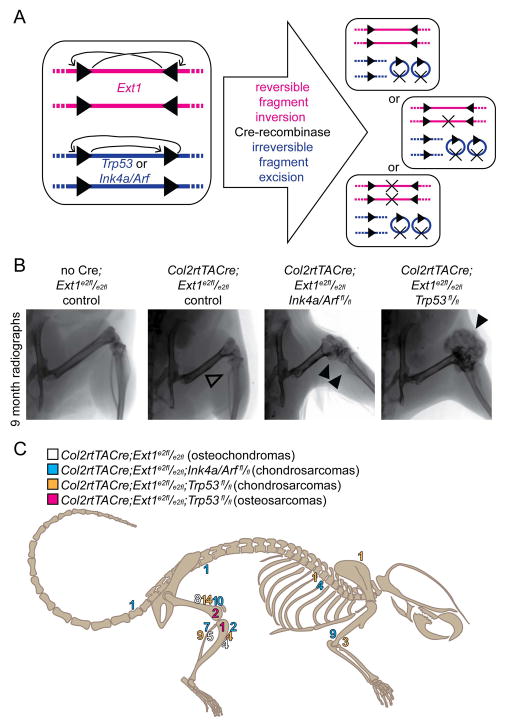
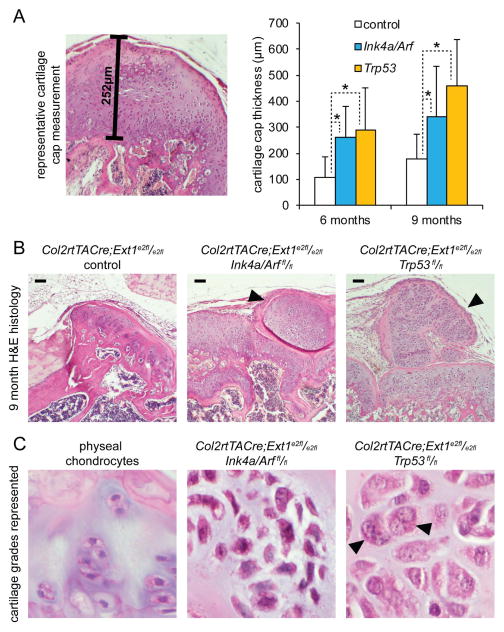
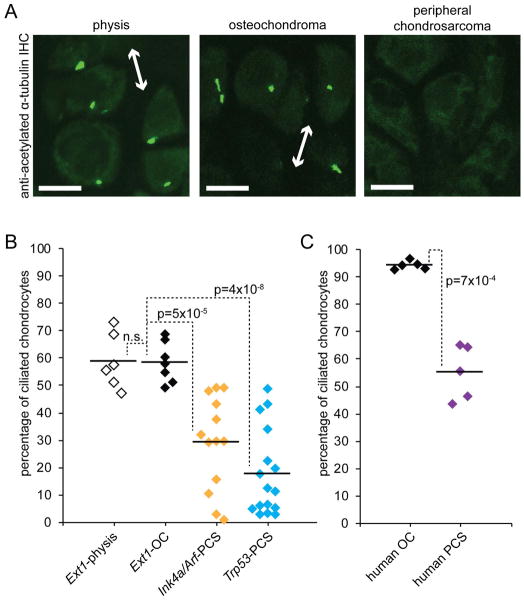
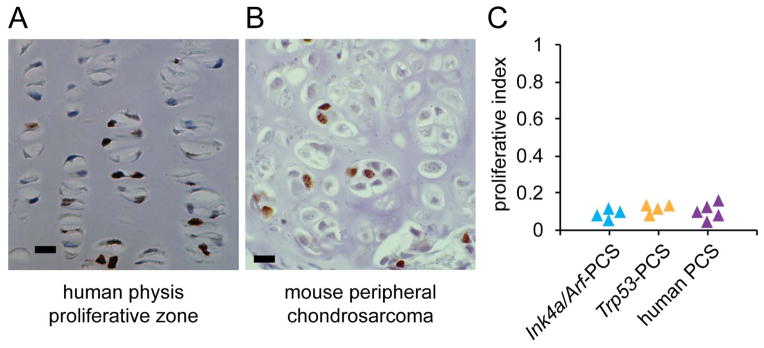
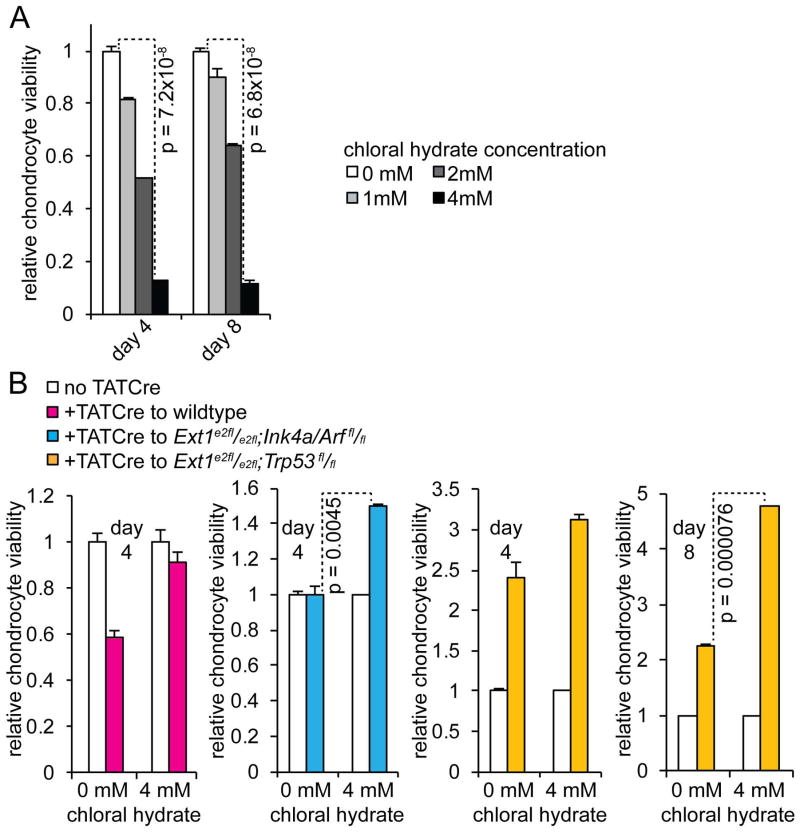
Peripheral chondrosarcoma (PCS) develops as malignant transformation of an osteochondroma, a benign cartilaginous outgrowth at the bone surface. Its invasive, lobular growth despite low-grade histology suggests a loss of chondrocyte polarity. The known genetics of osteochondromagenesis include mosaic loss of EXT1 or EXT2 in both hereditary and non-hereditary cases. The most frequent genetic aberrations in human PCS also include disruptions of CDKN2A or TP53. In order to test the sufficiency of either of these to drive progression of an osteochondroma to PCS, we added conditional loss of Trp53 or Ink4a/Arf in an Ext1-driven mouse model of osteochondromagenesis. Each additional tumour suppressor silencing efficiently drove the development of growths that mimic human PCS. As in humans, lobules developed from both Ext1-null and Ext1-functional clones within osteochondromas. Assessment of their orientation revealed an absence of primary cilia in the majority of mouse PCS chondrocytes, which was corroborated in human PCSs. Loss of primary cilia may be responsible for the lost polarity phenotype ascribed to PCS. Cilia deficiency blocks proliferation in physeal chondrocytes, but cell cycle deregulation is sufficient to rescue chondrocyte proliferation following deciliation. This provides a basis of selective pressure for the frequent cell-cycle regulator silencing observed in peripheral chondrosarcomagenesis. Mosaic loss of Ext1 combined with loss of cell cycle regulators promotes peripheral chondrosarcomagenesis in the mouse and reveals deficient ciliogenesis in both the model and the human disease, explaining biological behaviour including lobular and invasive growth.
Keywords: chondrosarcoma; mouse genetic model; osteochondroma; primary cilium; tumour suppressor.
Copyright © 2015 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
Conflict of interest statement
Figures
References
-
- Giuffrida AY, Burgueno JE, Koniaris LG, et al. Chondrosarcoma in the United States (1973 to 2003): an analysis of 2890 cases from the SEER database. J Bone Joint Surg Am. 2009;91:1063–1072. - PubMed
-
- Bovee JV, Cleton-Jansen AM, Kuipers-Dijkshoorn NJ, et al. Loss of heterozygosity and DNA ploidy point to a diverging genetic mechanism in the origin of peripheral and central chondrosarcoma. Genes Chromosomes Cancer. 1999;26:237–246. - PubMed
-
- Lin PP, Moussallem CD, Deavers MT. Secondary chondrosarcoma. J Am Acad Orthop Surg. 2010;18:608–615. - PubMed
-
- Ahmed AR, Tan TS, Unni KK, et al. Secondary chondrosarcoma in osteochondroma: report of 107 patients. Clin Orthop Relat Res. 2003:193–206. - PubMed
-
- Fletcher CDM World Health Organization. WHO classification of tumours of soft tissue and bone. 4. IARC Press; Lyon: 2013. International Agency for Research on Cancer.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
