

Probing the sources of the apparent irreproducibility of amyloid formation: drastic changes in kinetics and a switch in mechanism due to micellelike oligomer formation at critical concentrations of IAPP
- PMID: 25645610
- PMCID: PMC11444341
- DOI: 10.1021/jp511758w
Probing the sources of the apparent irreproducibility of amyloid formation: drastic changes in kinetics and a switch in mechanism due to micellelike oligomer formation at critical concentrations of IAPP
Abstract
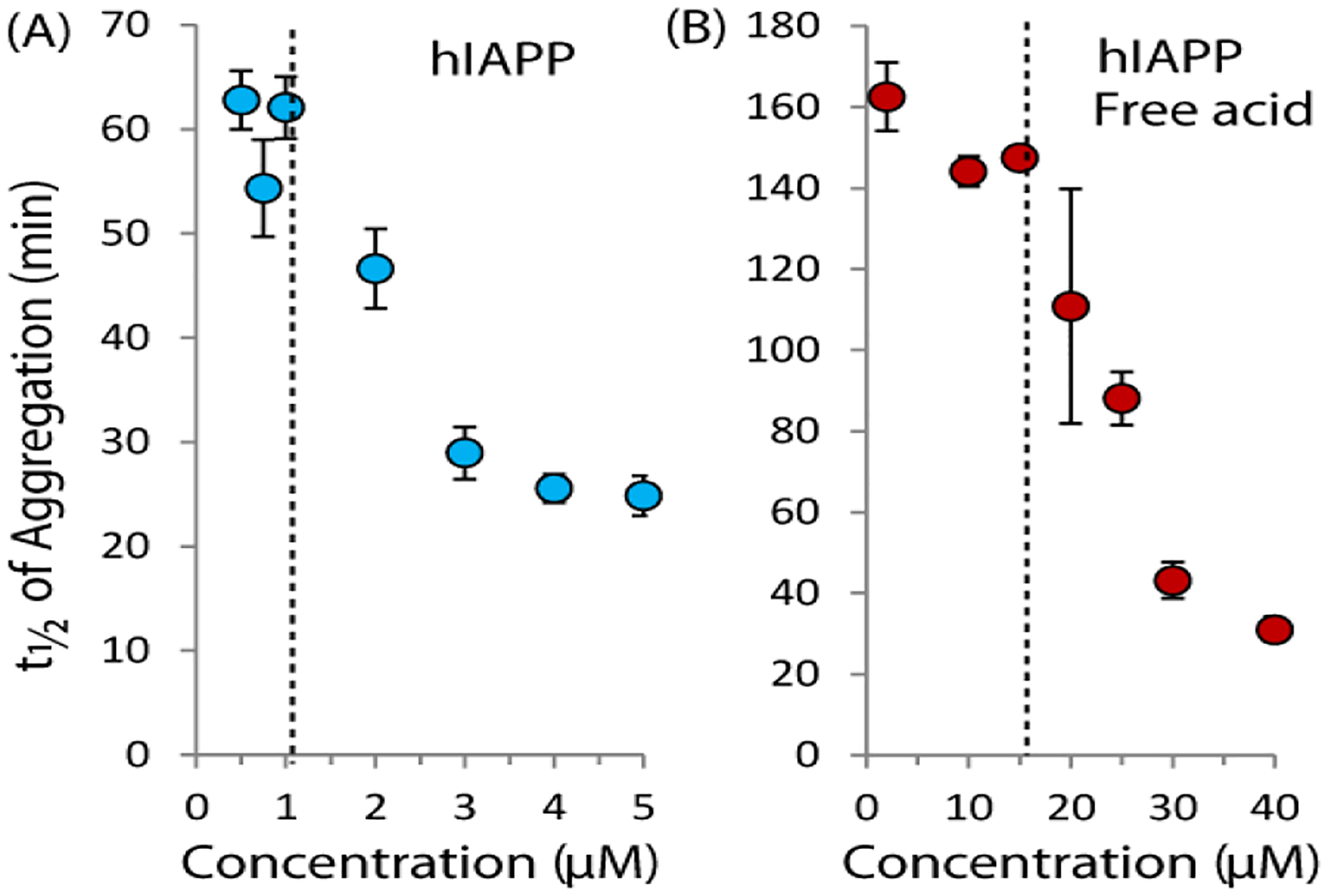
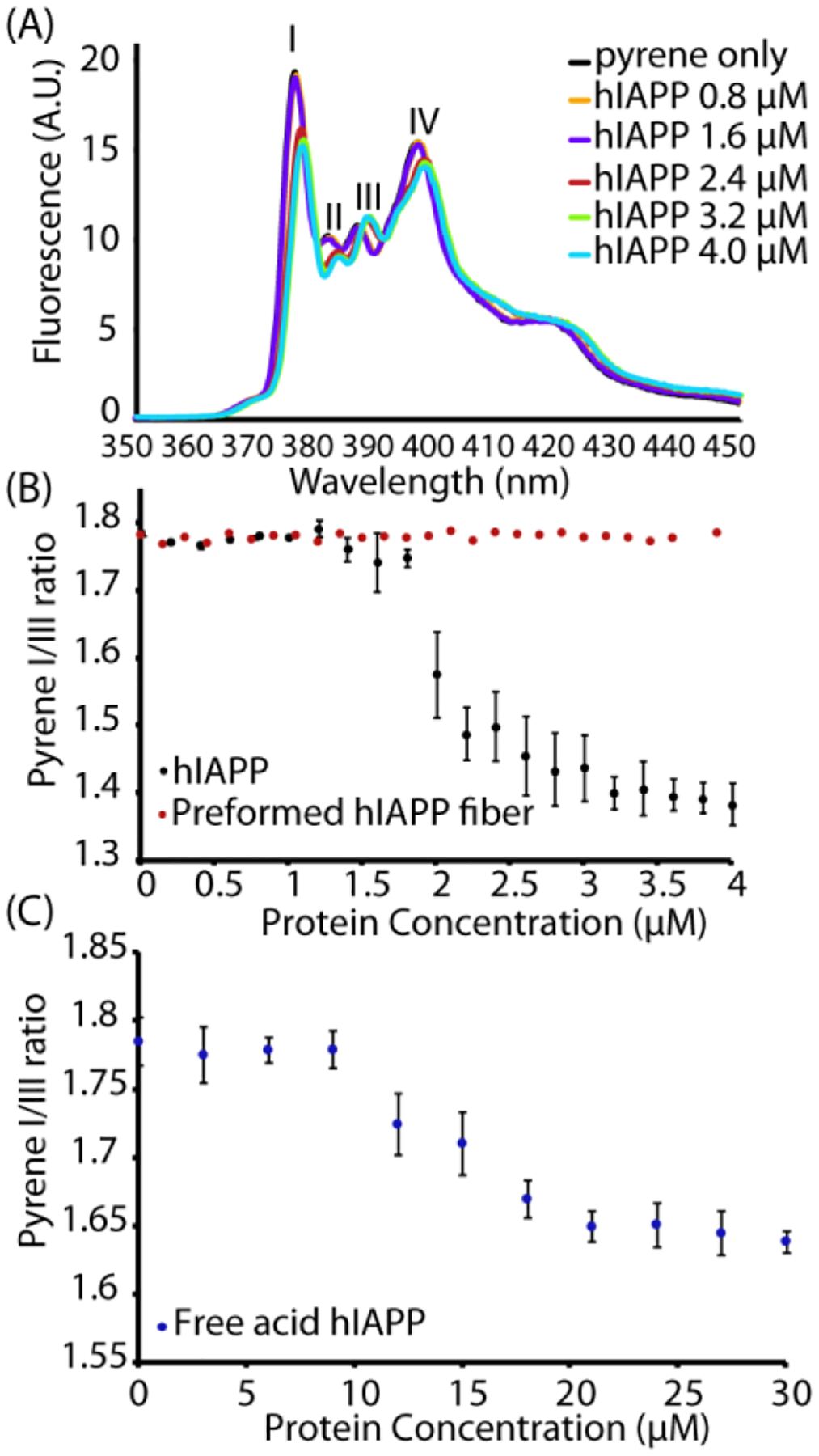
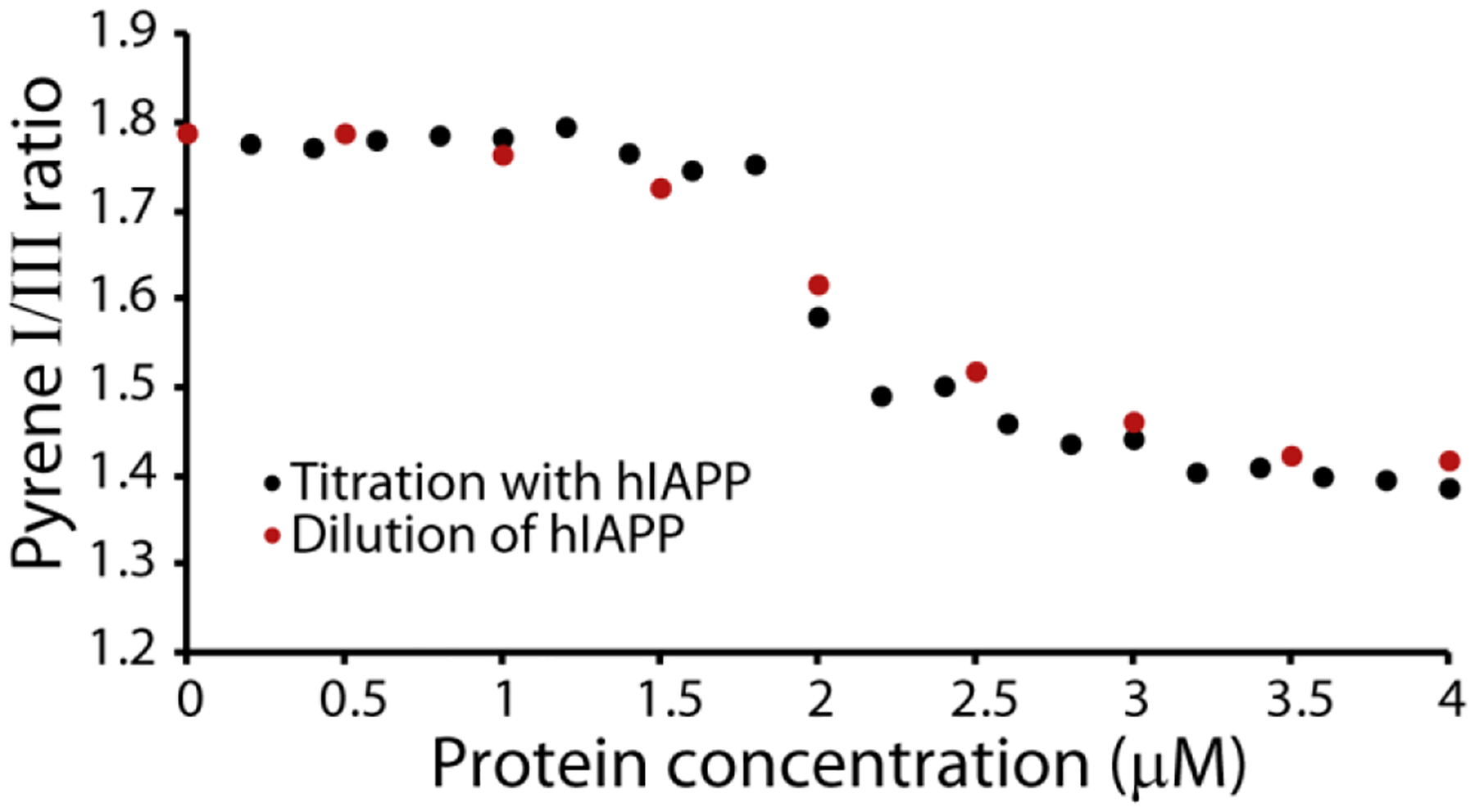
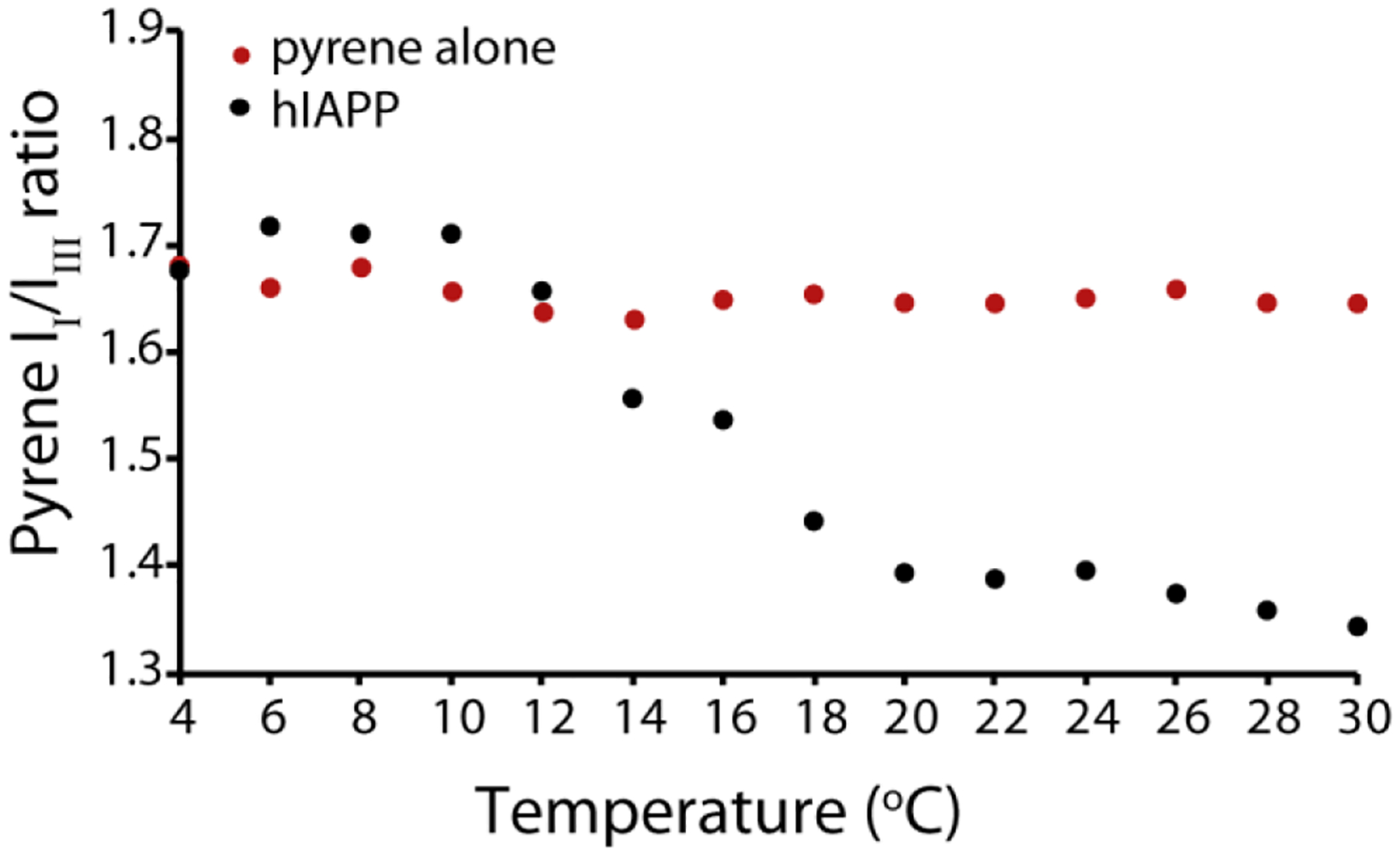
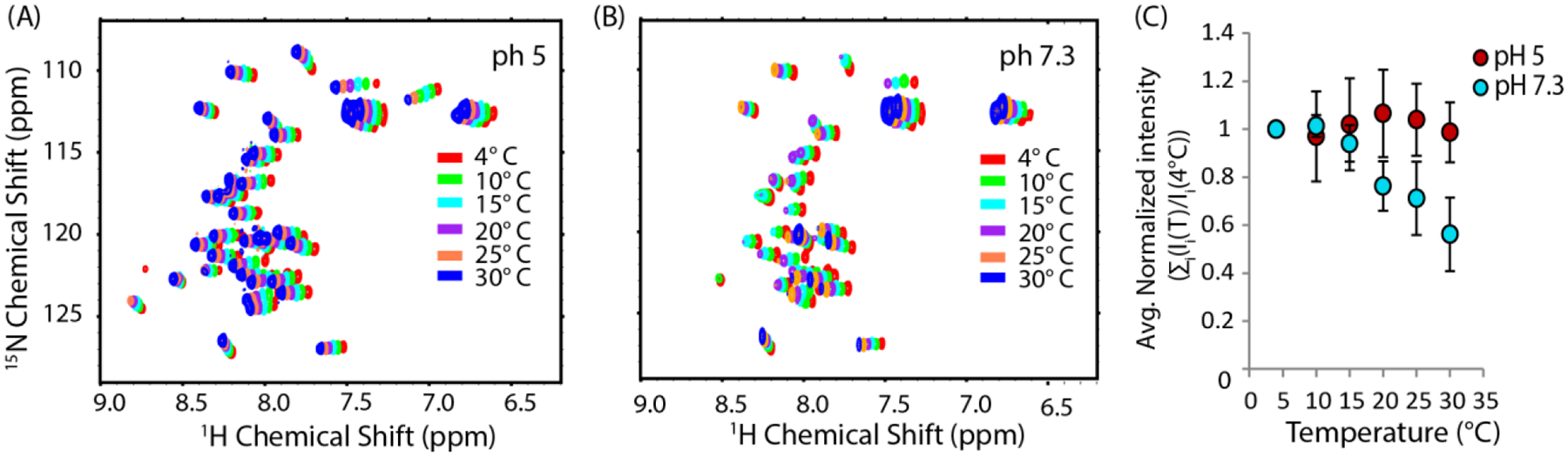
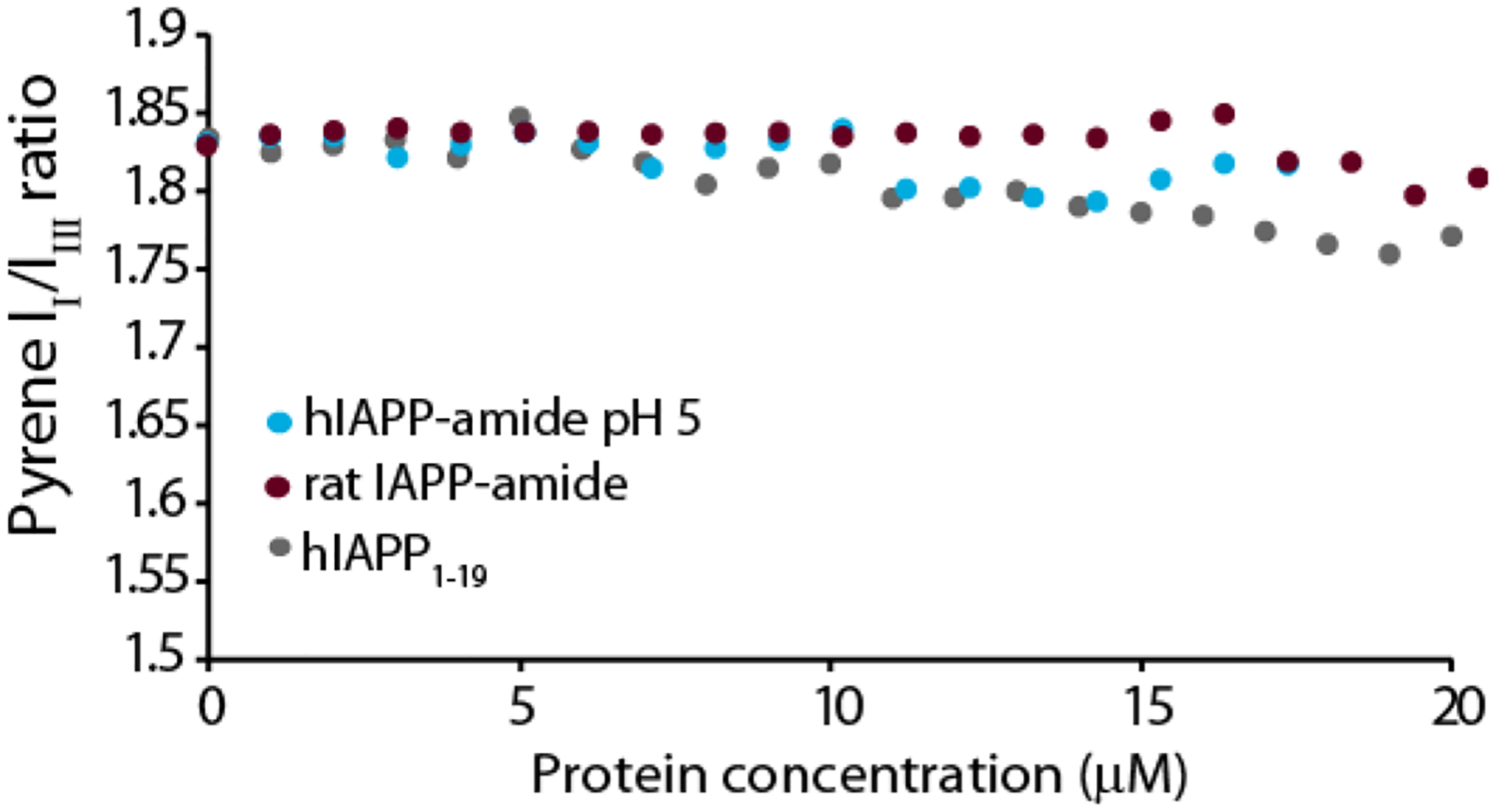
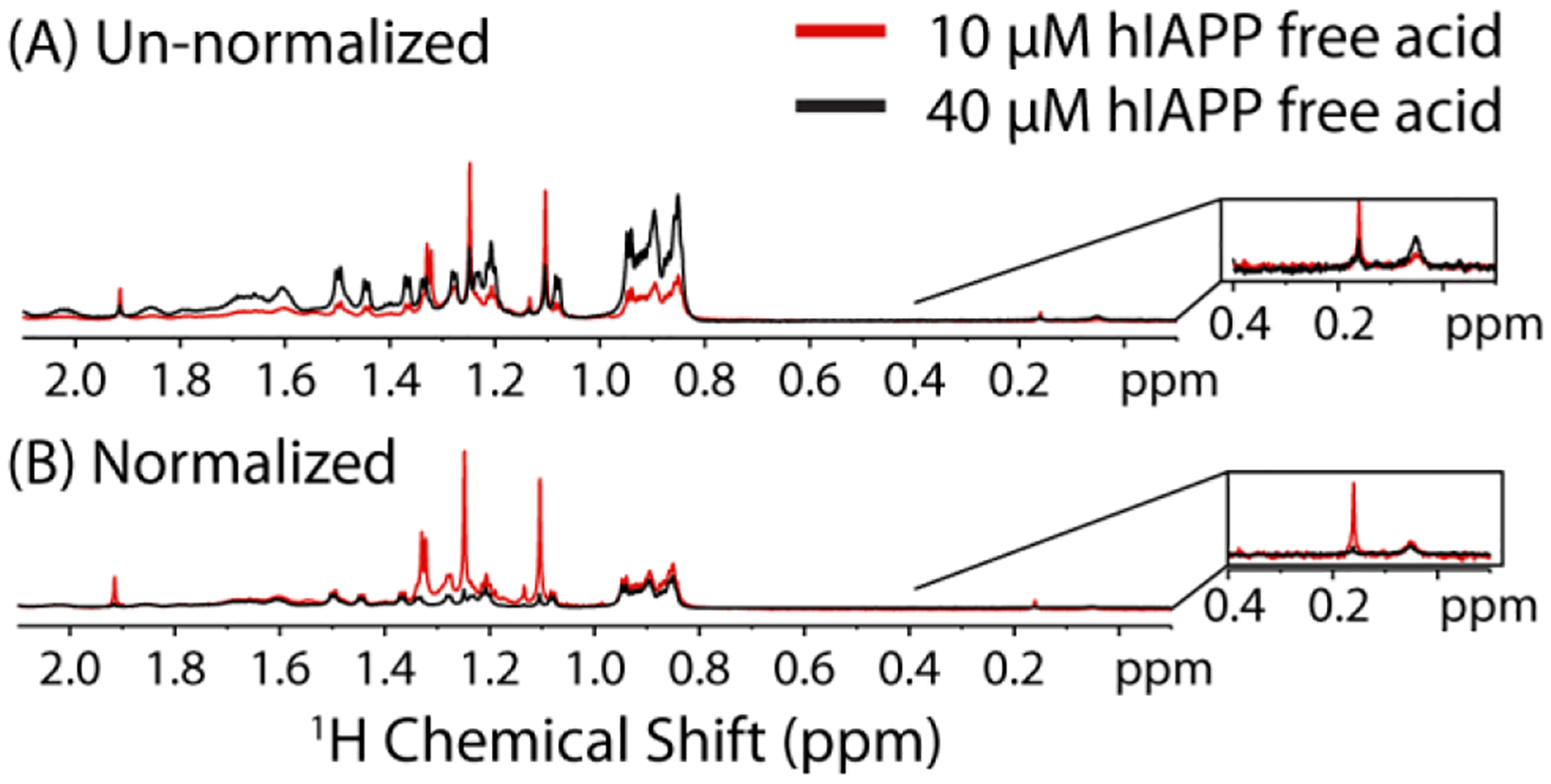
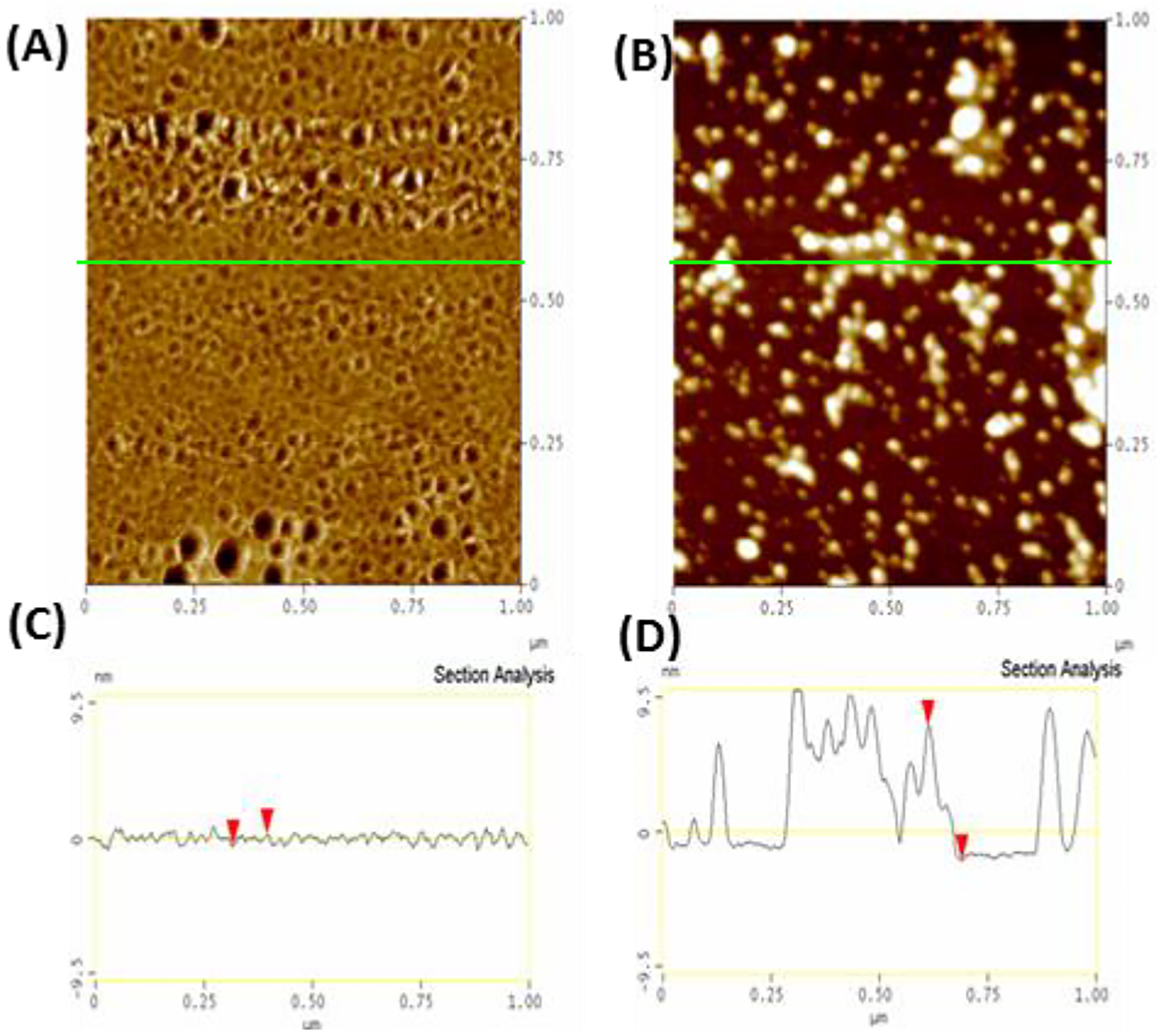
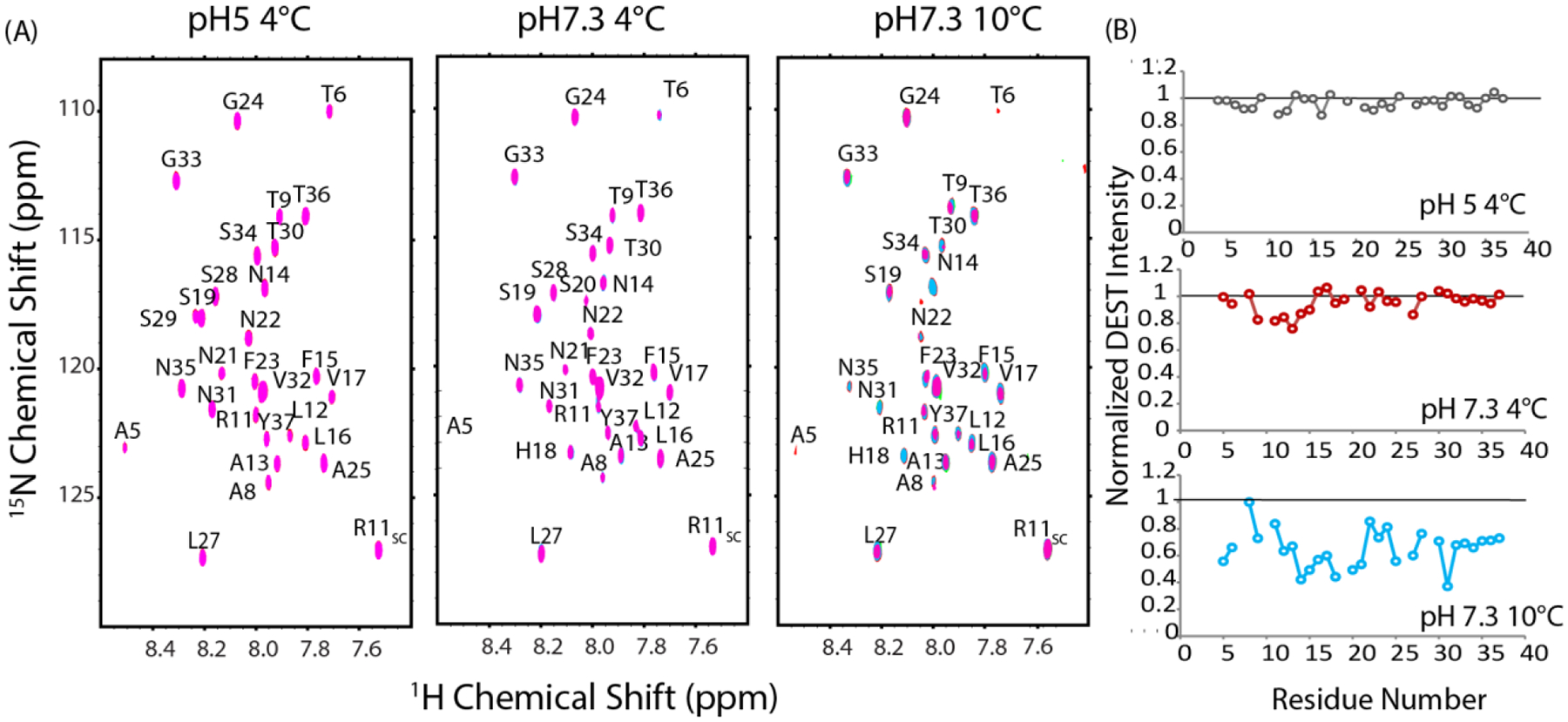
The aggregation of amyloidogenic proteins is infamous for being highly chaotic, with small variations in conditions sometimes leading to large changes in aggregation rates. Using the amyloidogenic protein IAPP (islet amyloid polypeptide protein, also known as amylin) as an example, we show that a part of this phenomenon may be related to the formation of micellelike oligomers at specific critical concentrations and temperatures. We show that pyrene fluorescence can sensitively detect micellelike oligomer formation by IAPP and discriminate between micellelike oligomers from fibers and monomers, making pyrene one of the few chemical probes specific to a prefibrillar oligomer. We further show that oligomers of this type reversibly form at critical concentrations in the low micromolar range and at specific critical temperatures. Micellelike oligomer formation has several consequences for amyloid formation by IAPP. First, the kinetics of fiber formation increase substantially as the critical concentration is approached but are nearly independent of concentration below it, suggesting a direct role for the oligomers in fiber formation. Second, the critical concentration is strongly correlated with the propensity to form amyloid: higher critical concentrations are observed for both IAPP variants with lower amyloidogenicity and for native IAPP at acidic pH in which aggregation is greatly slowed. Furthermore, using the DEST NMR technique, we show that the pathway of amyloid formation switches as the critical point is approached, with self-interactions primarily near the N-terminus below the critical temperature and near the central region above the critical temperature, reconciling two apparently conflicting views of the initiation of IAPP aggregation.
Figures
References
-
- Ross CA; Poirier MA, Protein Aggregation and Neurodegenerative Disease. Nat. Med 2004, 10, S10–S17. - PubMed
-
- Harrison RS; Sharpe PC; Singh Y; Fairlie DP, Amyloid Peptides and Proteins in Review. Rev. Physiol. Biochem. Pharm 2007, 159, 1–77. - PubMed
-
- Hamley IW, The Amyloid Beta Peptide: A Chemist’s Perspective. Role in Alzheimer’s and Fibrillization. Chem. Rev 2012, 112, 5147–5192. - PubMed
-
- Marchesi VT, Alzheimer’s Disease 2012: The Great Amyloid Gamble. Am. J. Pathol 2012, 180, 1762–1767. - PubMed
-
- Liu TY; Bitan G, Modulating Self-Assembly of Amyloidogenic Proteins as a Therapeutic Approach for Neurodegenerative Diseases: Strategies and Mechanisms. ChemMedChem 2012, 7, 359–374. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
