

Dense fibrillar collagen is a potent inducer of invadopodia via a specific signaling network
- PMID: 25646088
- PMCID: PMC4315243
- DOI: 10.1083/jcb.201405099
Dense fibrillar collagen is a potent inducer of invadopodia via a specific signaling network
Abstract
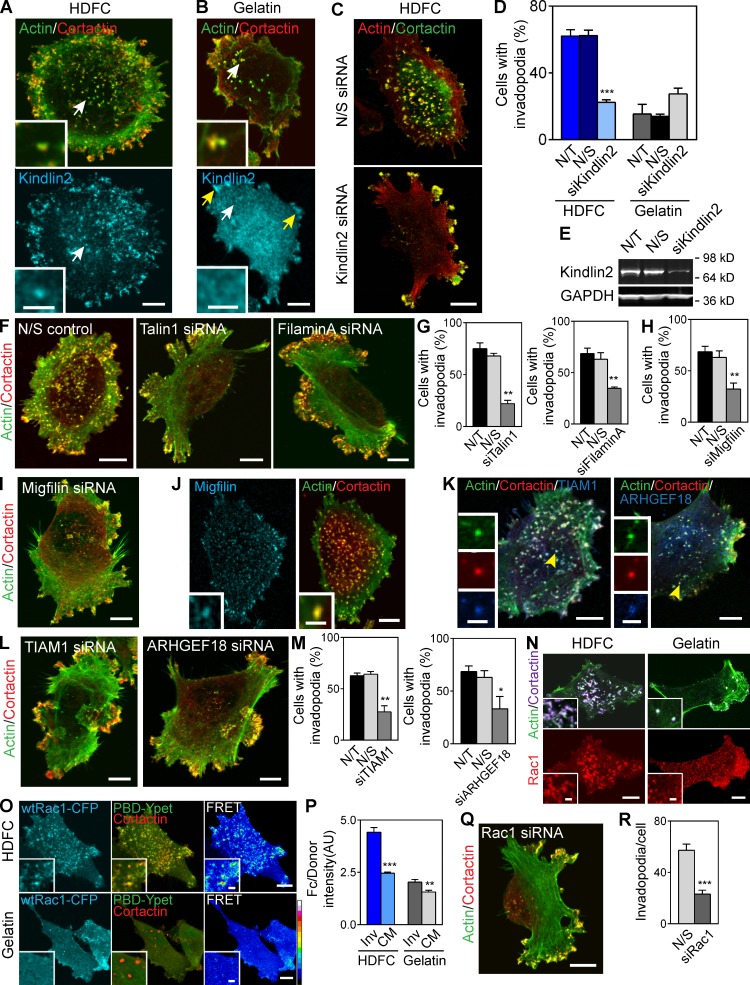
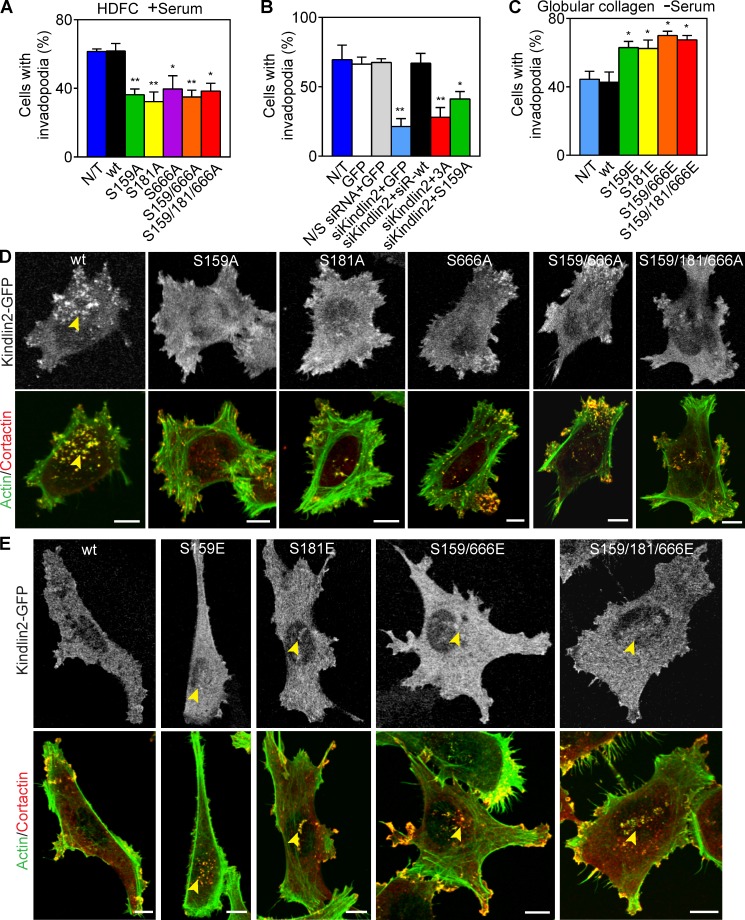
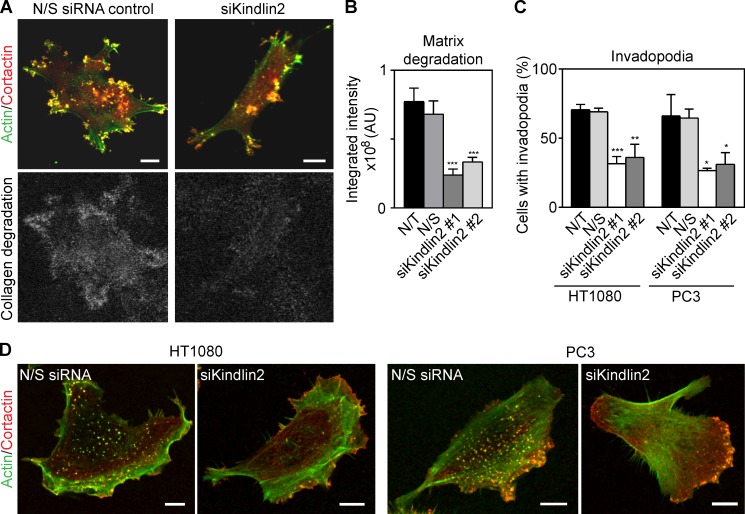
Cell interactions with the extracellular matrix (ECM) can regulate multiple cellular activities and the matrix itself in dynamic, bidirectional processes. One such process is local proteolytic modification of the ECM. Invadopodia of tumor cells are actin-rich proteolytic protrusions that locally degrade matrix molecules and mediate invasion. We report that a novel high-density fibrillar collagen (HDFC) matrix is a potent inducer of invadopodia, both in carcinoma cell lines and in primary human fibroblasts. In carcinoma cells, HDFC matrix induced formation of invadopodia via a specific integrin signaling pathway that did not require growth factors or even altered gene and protein expression. In contrast, phosphoproteomics identified major changes in a complex phosphosignaling network with kindlin2 serine phosphorylation as a key regulatory element. This kindlin2-dependent signal transduction network was required for efficient induction of invadopodia on dense fibrillar collagen and for local degradation of collagen. This novel phosphosignaling mechanism regulates cell surface invadopodia via kindlin2 for local proteolytic remodeling of the ECM.
Figures
Similar articles
-
Dependence of invadopodia function on collagen fiber spacing and cross-linking: computational modeling and experimental evidence.Biophys J. 2008 Sep;95(5):2203-18. doi: 10.1529/biophysj.108.133199. Epub 2008 May 30. Biophys J. 2008. PMID: 18515372 Free PMC article.
-
The interplay between the proteolytic, invasive, and adhesive domains of invadopodia and their roles in cancer invasion.Cell Adh Migr. 2014;8(3):215-25. doi: 10.4161/cam.27842. Cell Adh Migr. 2014. PMID: 24714132 Free PMC article. Review.
-
Preparation of High-Density Fibrillar Collagen Matrices That Mimic Desmoplastic Tumor Stroma.Curr Protoc Cell Biol. 2016 Mar 1;70:10.19.1-10.19.11. doi: 10.1002/0471143030.cb1019s70. Curr Protoc Cell Biol. 2016. PMID: 26930554 Free PMC article.
-
Pathological roles of invadopodia in cancer invasion and metastasis.Eur J Cell Biol. 2012 Nov-Dec;91(11-12):902-7. doi: 10.1016/j.ejcb.2012.04.005. Epub 2012 Jun 2. Eur J Cell Biol. 2012. PMID: 22658792 Review.
-
Extracellular matrix rigidity promotes invadopodia activity.Curr Biol. 2008 Sep 9;18(17):1295-1299. doi: 10.1016/j.cub.2008.07.090. Epub 2008 Aug 21. Curr Biol. 2008. PMID: 18718759 Free PMC article.
Cited by
-
A computational modeling of invadopodia protrusion into an extracellular matrix fiber network.Sci Rep. 2022 Jan 24;12(1):1231. doi: 10.1038/s41598-022-05224-9. Sci Rep. 2022. PMID: 35075179 Free PMC article.
-
SRC and TKS5 mediated podosome formation in fibroblasts promotes extracellular matrix invasion and pulmonary fibrosis.Nat Commun. 2023 Sep 21;14(1):5882. doi: 10.1038/s41467-023-41614-x. Nat Commun. 2023. PMID: 37735172 Free PMC article.
-
IKKε phosphorylates kindlin-2 to induce invadopodia formation and promote colorectal cancer metastasis.Theranostics. 2020 Jan 16;10(5):2358-2373. doi: 10.7150/thno.40397. eCollection 2020. Theranostics. 2020. PMID: 32104508 Free PMC article.
-
Matrix rigidity differentially regulates invadopodia activity through ROCK1 and ROCK2.Biomaterials. 2016 Apr;84:119-129. doi: 10.1016/j.biomaterials.2016.01.028. Epub 2016 Jan 15. Biomaterials. 2016. PMID: 26826790 Free PMC article.
-
Basement Membranes in Development and Disease.Curr Top Dev Biol. 2018;130:143-191. doi: 10.1016/bs.ctdb.2018.02.005. Epub 2018 Mar 31. Curr Top Dev Biol. 2018. PMID: 29853176 Free PMC article. Review.
References
-
- Artym V.V., Zhang Y., Seillier-Moiseiwitsch F., Yamada K.M., and Mueller S.C.. 2006. Dynamic interactions of cortactin and membrane type 1 matrix metalloproteinase at invadopodia: defining the stages of invadopodia formation and function. Cancer Res. 66:3034–3043 10.1158/0008-5472.CAN-05-2177 - DOI - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
