

Mitochondrial peroxiredoxin functions as crucial chaperone reservoir in Leishmania infantum
- PMID: 25646478
- PMCID: PMC4343147
- DOI: 10.1073/pnas.1419682112
Mitochondrial peroxiredoxin functions as crucial chaperone reservoir in Leishmania infantum
Abstract
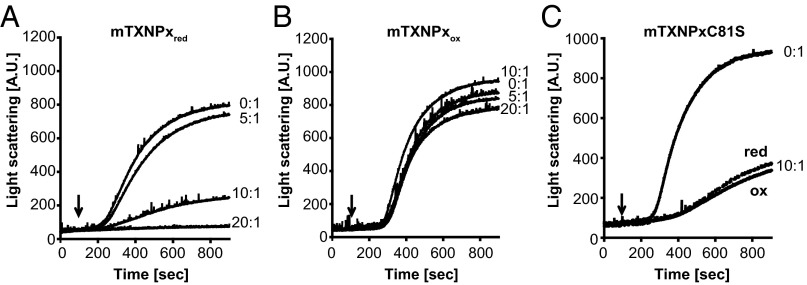
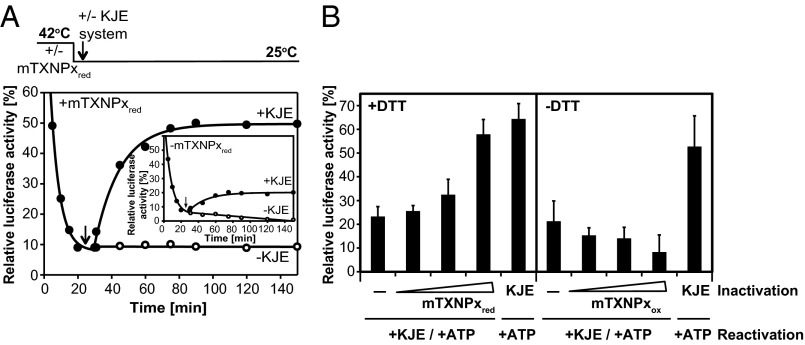
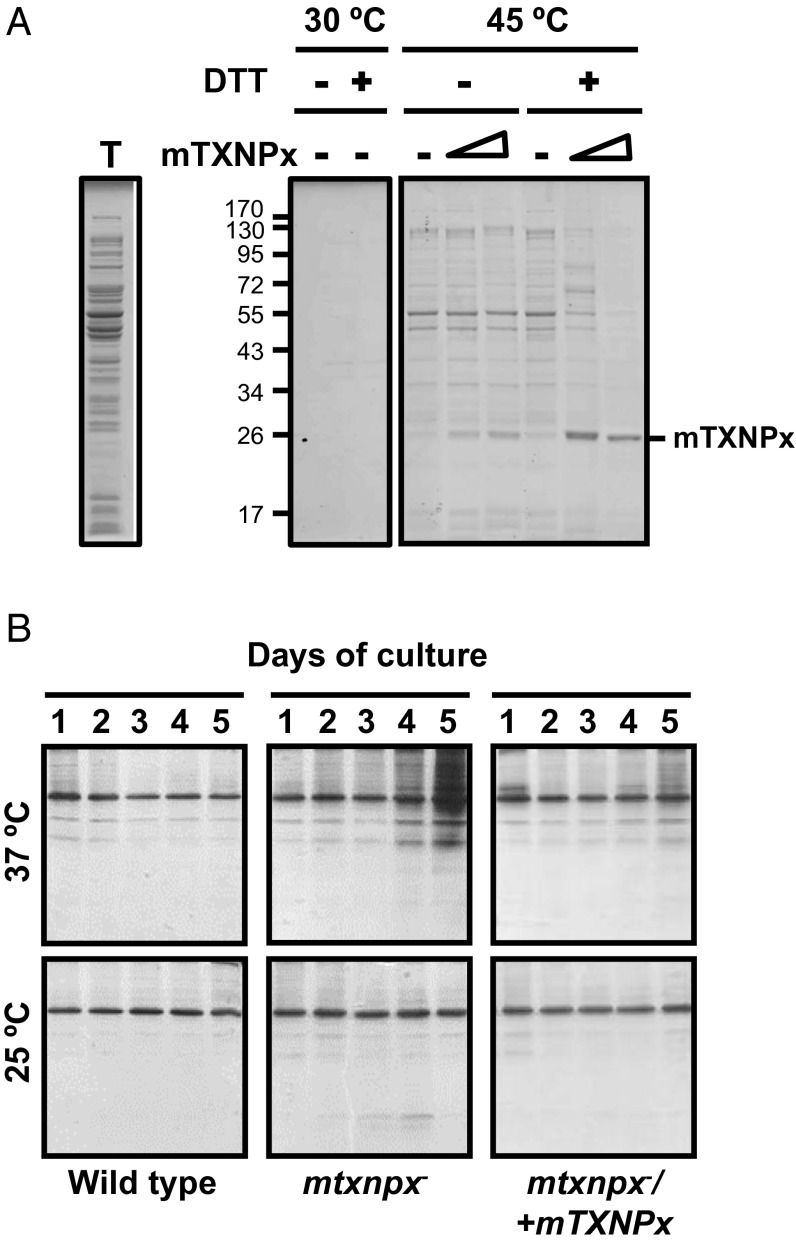
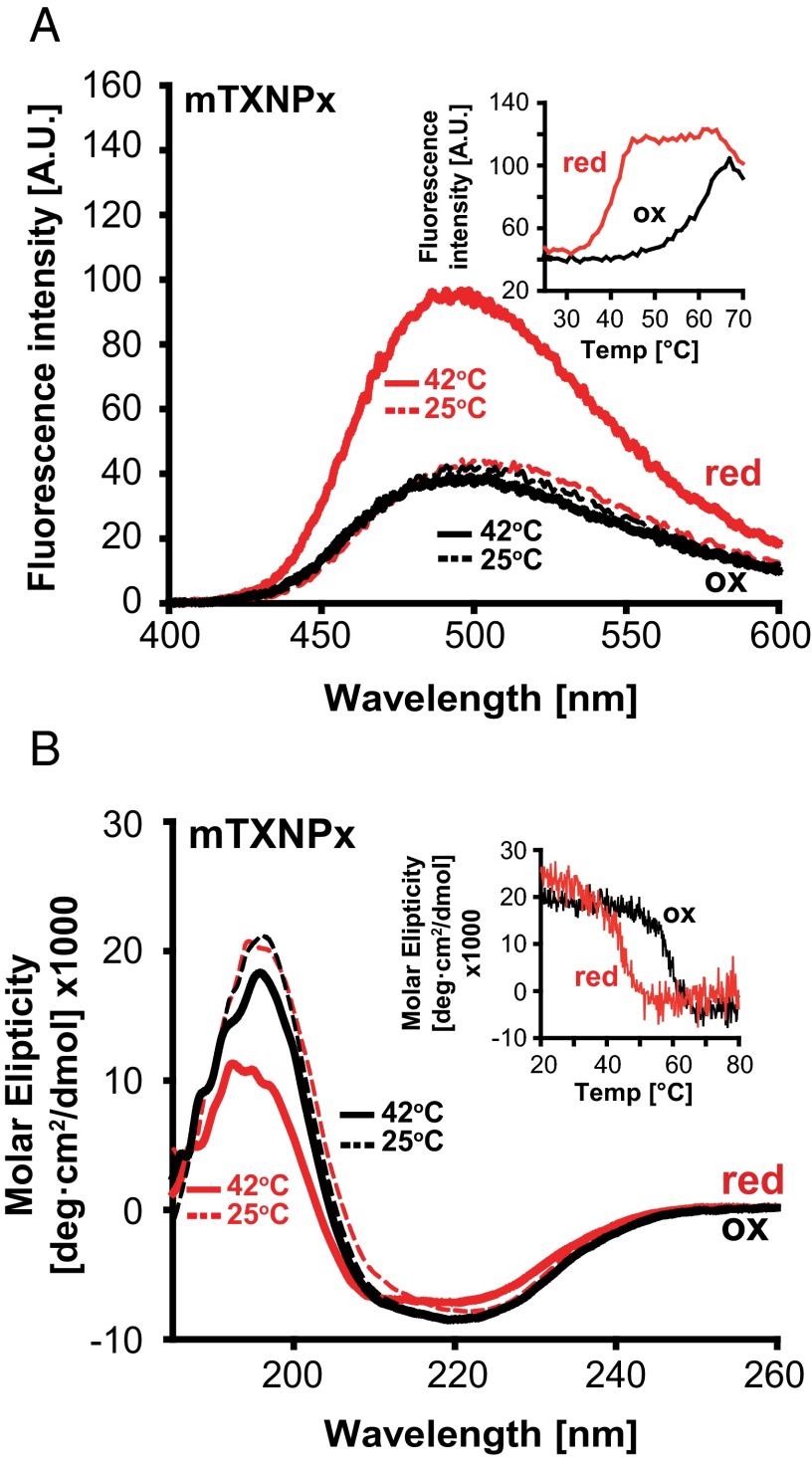
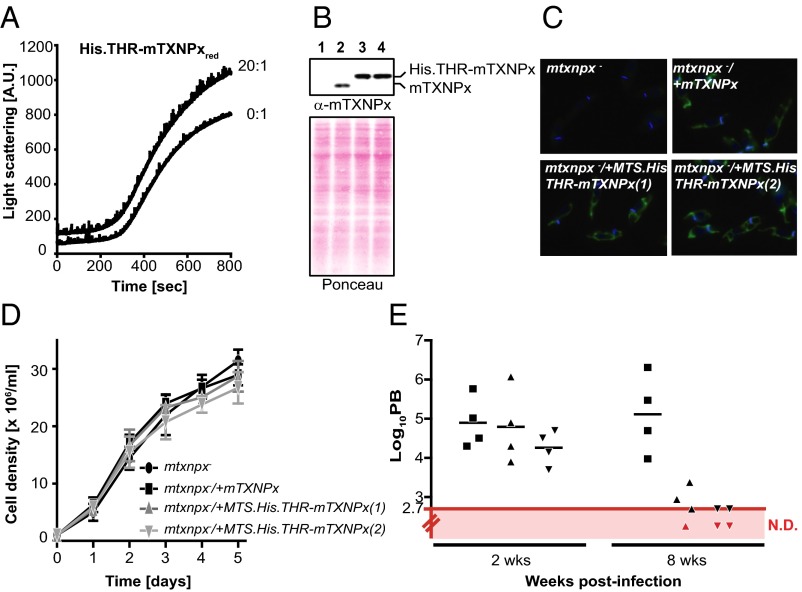
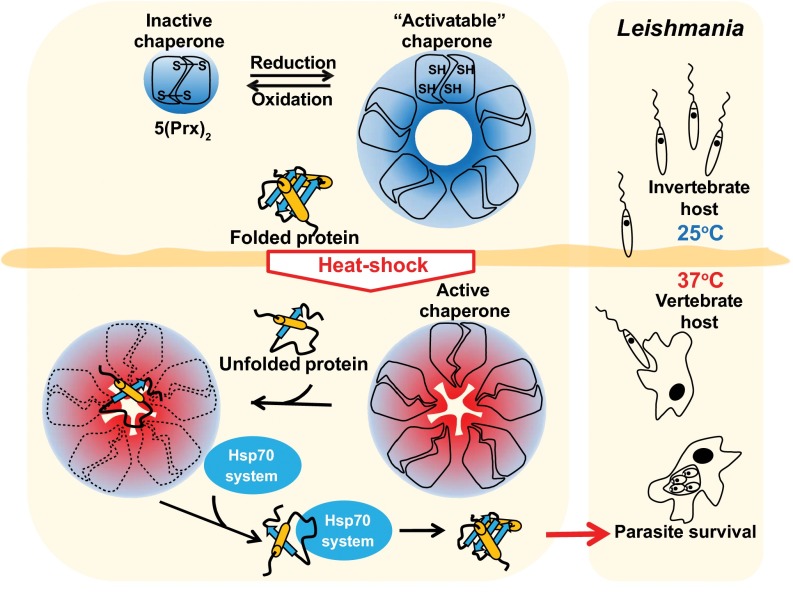
Cytosolic eukaryotic 2-Cys-peroxiredoxins have been widely reported to act as dual-function proteins, either detoxifying reactive oxygen species or acting as chaperones to prevent protein aggregation. Several stimuli, including peroxide-mediated sulfinic acid formation at the active site cysteine, have been proposed to trigger the chaperone activity. However, the mechanism underlying this activation and the extent to which the chaperone function is crucial under physiological conditions in vivo remained unknown. Here we demonstrate that in the vector-borne protozoan parasite Leishmania infantum, mitochondrial peroxiredoxin (Prx) exerts intrinsic ATP-independent chaperone activity, protecting a wide variety of different proteins against heat stress-mediated unfolding in vitro and in vivo. Activation of the chaperone function appears to be induced by temperature-mediated restructuring of the reduced decamers, promoting binding of unfolding client proteins in the center of Prx's ringlike structure. Client proteins are maintained in a folding-competent conformation until restoration of nonstress conditions, upon which they are released and transferred to ATP-dependent chaperones for refolding. Interference with client binding impairs parasite infectivity, providing compelling evidence for the in vivo importance of Prx's chaperone function. Our results suggest that reduced Prx provides a mitochondrial chaperone reservoir, which allows L. infantum to deal successfully with protein unfolding conditions during the transition from insect to the mammalian hosts and to generate viable parasites capable of perpetuating infection.
Keywords: Leishmania; chaperone; peroxiredoxin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Chae HZ, Chung SJ, Rhee SG. Thioredoxin-dependent peroxide reductase from yeast. J Biol Chem. 1994;269(44):27670–27678. - PubMed
-
- Hofmann B, Hecht HJ, Flohé L. Peroxiredoxins. Biol Chem. 2002;383(3-4):347–364. - PubMed
-
- Rhee SG, Woo HA. Multiple functions of peroxiredoxins: peroxidases, sensors and regulators of the intracellular messenger H2O2, and protein chaperones. Antioxid Redox Signaling. 2011;15(3):781–794. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
