

Crystal structures of the carboxyl cGMP binding domain of the Plasmodium falciparum cGMP-dependent protein kinase reveal a novel capping triad crucial for merozoite egress
- PMID: 25646845
- PMCID: PMC4412288
- DOI: 10.1371/journal.ppat.1004639
Crystal structures of the carboxyl cGMP binding domain of the Plasmodium falciparum cGMP-dependent protein kinase reveal a novel capping triad crucial for merozoite egress
Abstract
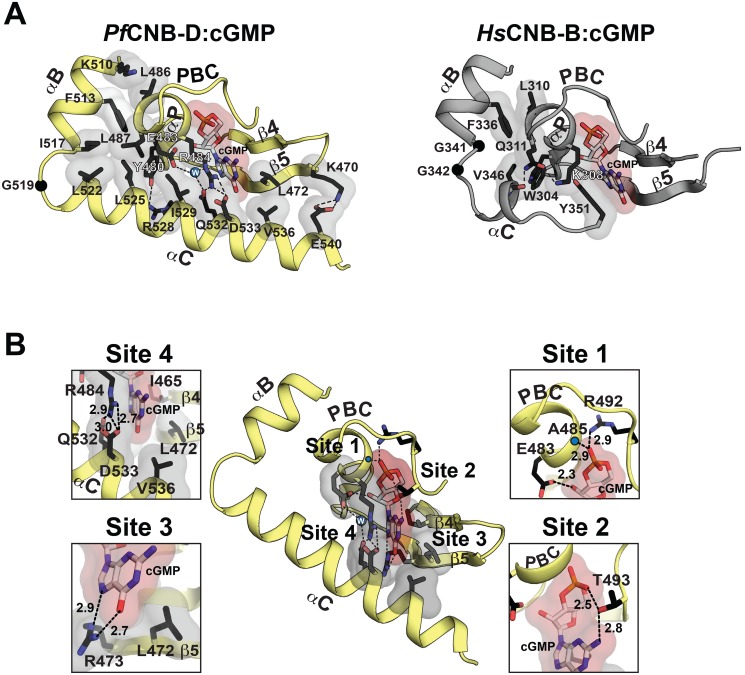
The Plasmodium falciparum cGMP-dependent protein kinase (PfPKG) is a key regulator across the malaria parasite life cycle. Little is known about PfPKG's activation mechanism. Here we report that the carboxyl cyclic nucleotide binding domain functions as a "gatekeeper" for activation by providing the highest cGMP affinity and selectivity. To understand the mechanism, we have solved its crystal structures with and without cGMP at 2.0 and 1.9 Å, respectively. These structures revealed a PfPKG-specific capping triad that forms upon cGMP binding, and disrupting the triad reduces kinase activity by 90%. Furthermore, mutating these residues in the parasite prevents blood stage merozoite egress, confirming the essential nature of the triad in the parasite. We propose a mechanism of activation where cGMP binding allosterically triggers the conformational change at the αC-helix, which bridges the regulatory and catalytic domains, causing the capping triad to form and stabilize the active conformation.
Conflict of interest statement
The authors have declared that no competing interests exist. The authors also have no affiliation or competing interest with the Eli Lilly company. Eli Lilly has no role in designing and conducting this study. This does not alter our adherence to all PLOS Pathogens policies on sharing data and materials.
Figures




References
-
- Cowman AF, Crabb BS (2006) Invasion of red blood cells by malaria parasites. Cell 124: 755–766. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
