

Exploring bacterial organelle interactomes: a model of the protein-protein interaction network in the Pdu microcompartment
- PMID: 25646976
- PMCID: PMC4315436
- DOI: 10.1371/journal.pcbi.1004067
Exploring bacterial organelle interactomes: a model of the protein-protein interaction network in the Pdu microcompartment
Abstract
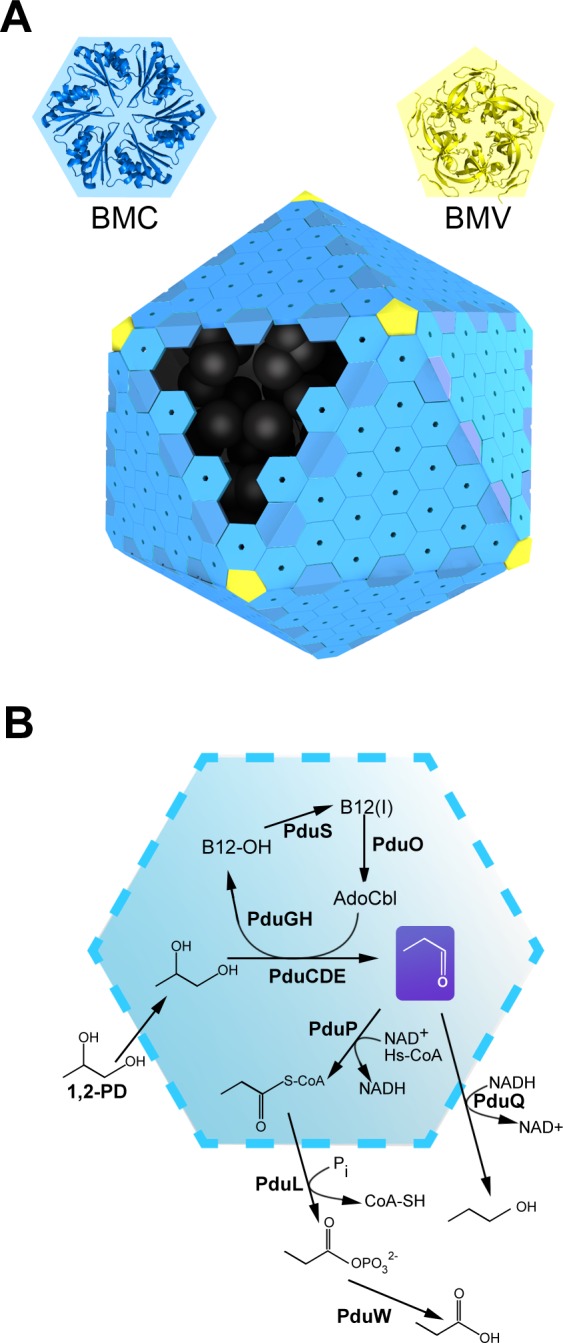
Bacterial microcompartments (MCPs) are protein-bound organelles that carry out diverse metabolic pathways in a wide range of bacteria. These supramolecular assemblies consist of a thin outer protein shell, reminiscent of a viral capsid, which encapsulates sequentially acting enzymes. The most complex MCP elucidated so far is the propanediol utilizing (Pdu) microcompartment. It contains the reactions for degrading 1,2-propanediol. While several experimental studies on the Pdu system have provided hints about its organization, a clear picture of how all the individual components interact has not emerged yet. Here we use co-evolution-based methods, involving pairwise comparisons of protein phylogenetic trees, to predict the protein-protein interaction (PPI) network governing the assembly of the Pdu MCP. We propose a model of the Pdu interactome, from which selected PPIs are further inspected via computational docking simulations. We find that shell protein PduA is able to serve as a "universal hub" for targeting an array of enzymes presenting special N-terminal extensions, namely PduC, D, E, L and P. The varied N-terminal peptides are predicted to bind in the same cleft on the presumptive luminal face of the PduA hexamer. We also propose that PduV, a protein of unknown function with remote homology to the Ras-like GTPase superfamily, is likely to localize outside the MCP, interacting with the protruding β-barrel of the hexameric PduU shell protein. Preliminary experiments involving a bacterial two-hybrid assay are presented that corroborate the existence of a PduU-PduV interaction. This first systematic computational study aimed at characterizing the interactome of a bacterial microcompartment provides fresh insight into the organization of the Pdu MCP.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures





Similar articles
-
In Salmonella enterica, Ethanolamine Utilization Is Repressed by 1,2-Propanediol To Prevent Detrimental Mixing of Components of Two Different Bacterial Microcompartments.J Bacteriol. 2015 Jul;197(14):2412-21. doi: 10.1128/JB.00215-15. Epub 2015 May 11. J Bacteriol. 2015. PMID: 25962913 Free PMC article.
-
Linking the Salmonella enterica 1,2-Propanediol Utilization Bacterial Microcompartment Shell to the Enzymatic Core via the Shell Protein PduB.J Bacteriol. 2022 Sep 20;204(9):e0057621. doi: 10.1128/jb.00576-21. Epub 2022 May 16. J Bacteriol. 2022. PMID: 35575582 Free PMC article.
-
The N Terminus of the PduB Protein Binds the Protein Shell of the Pdu Microcompartment to Its Enzymatic Core.J Bacteriol. 2017 Mar 28;199(8):e00785-16. doi: 10.1128/JB.00785-16. Print 2017 Apr 15. J Bacteriol. 2017. PMID: 28138097 Free PMC article.
-
Bacterial microcompartment organelles: protein shell structure and evolution.Annu Rev Biophys. 2010;39:185-205. doi: 10.1146/annurev.biophys.093008.131418. Annu Rev Biophys. 2010. PMID: 20192762 Free PMC article. Review.
-
The protein shells of bacterial microcompartment organelles.Curr Opin Struct Biol. 2011 Apr;21(2):223-31. doi: 10.1016/j.sbi.2011.01.006. Curr Opin Struct Biol. 2011. PMID: 21315581 Free PMC article. Review.
Cited by
-
Decoding the stoichiometric composition and organisation of bacterial metabolosomes.Nat Commun. 2020 Apr 24;11(1):1976. doi: 10.1038/s41467-020-15888-4. Nat Commun. 2020. PMID: 32332738 Free PMC article.
-
Engineering formation of multiple recombinant Eut protein nanocompartments in E. coli.Sci Rep. 2016 Apr 11;6:24359. doi: 10.1038/srep24359. Sci Rep. 2016. PMID: 27063436 Free PMC article.
-
Introducing noncanonical amino acids for studying and engineering bacterial microcompartments.Curr Opin Microbiol. 2021 Jun;61:67-72. doi: 10.1016/j.mib.2021.03.004. Epub 2021 Apr 1. Curr Opin Microbiol. 2021. PMID: 33813159 Free PMC article. Review.
-
Chemical probing provides insight into the native assembly state of a bacterial microcompartment.Structure. 2022 Apr 7;30(4):537-550.e5. doi: 10.1016/j.str.2022.02.002. Epub 2022 Feb 24. Structure. 2022. PMID: 35216657 Free PMC article.
-
Bacterial microcompartments.Nat Rev Microbiol. 2018 May;16(5):277-290. doi: 10.1038/nrmicro.2018.10. Epub 2018 Mar 5. Nat Rev Microbiol. 2018. PMID: 29503457 Free PMC article. Review.
References
-
- Mullineaux CW (1999) The thylakoid membranes of cyanobacteria: structure, dynamics and function. Funct Plant Biol 26: 671–677.
-
- Löwe J, Amos LA (1998) Crystal structure of the bacterial cell-division protein FtsZ. Nature 391: 203–206. - PubMed
-
- Van den Ent F, Amos LA, Löwe J (2001) Prokaryotic origin of the actin cytoskeleton. Nature 413: 39–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
