

Inflammation-induced cell proliferation potentiates DNA damage-induced mutations in vivo
- PMID: 25647331
- PMCID: PMC4372043
- DOI: 10.1371/journal.pgen.1004901
Inflammation-induced cell proliferation potentiates DNA damage-induced mutations in vivo
Abstract
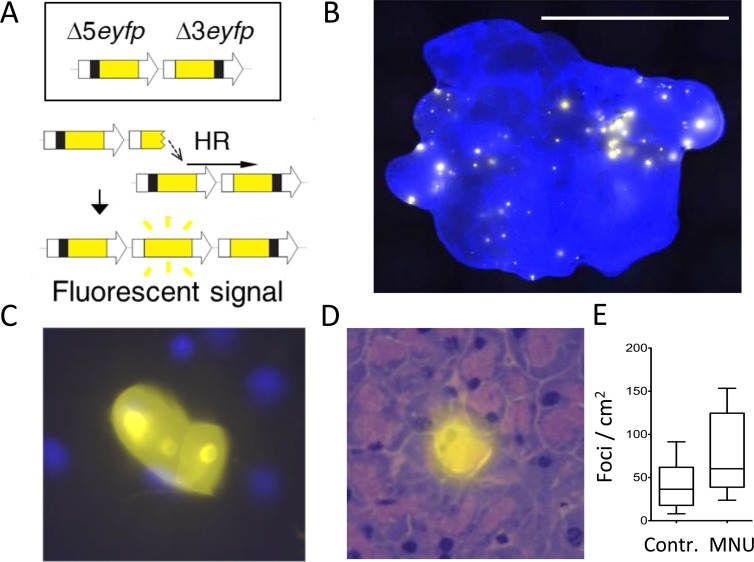
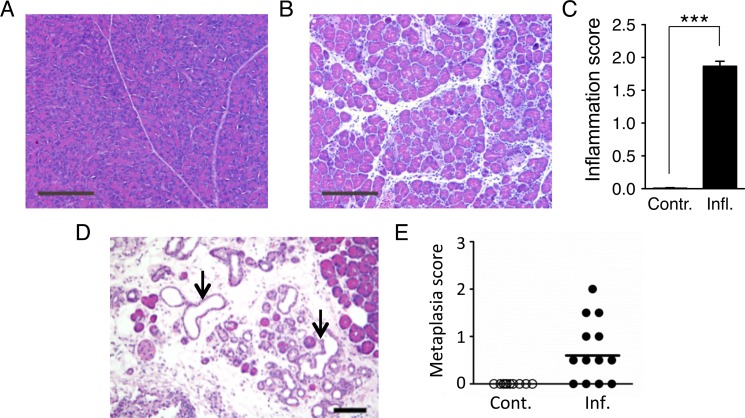
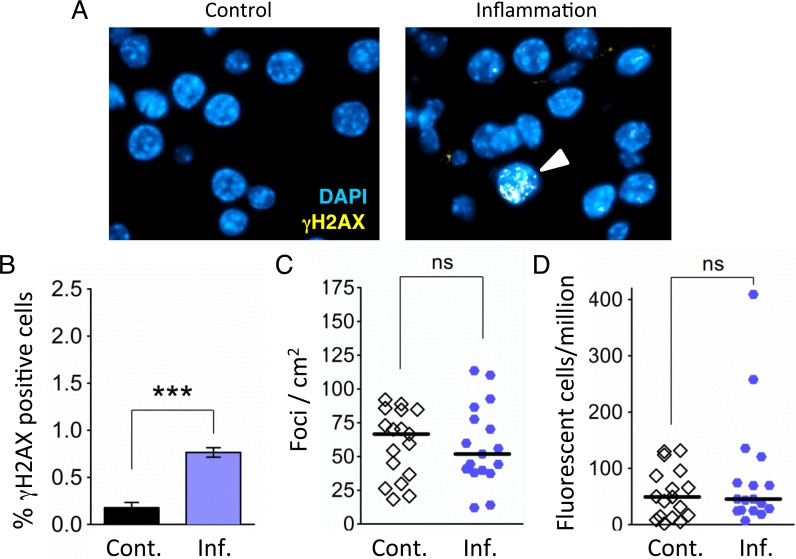
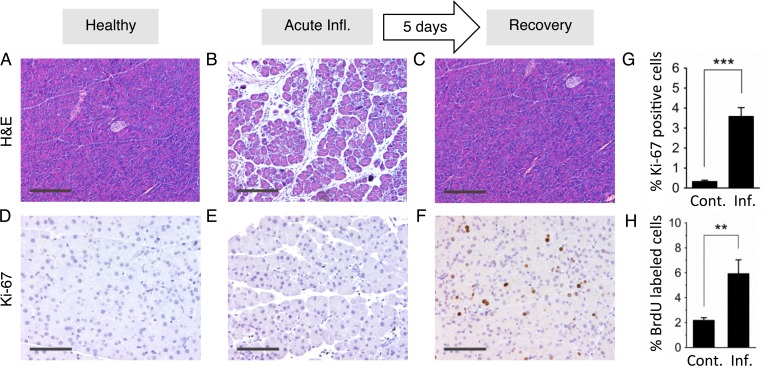
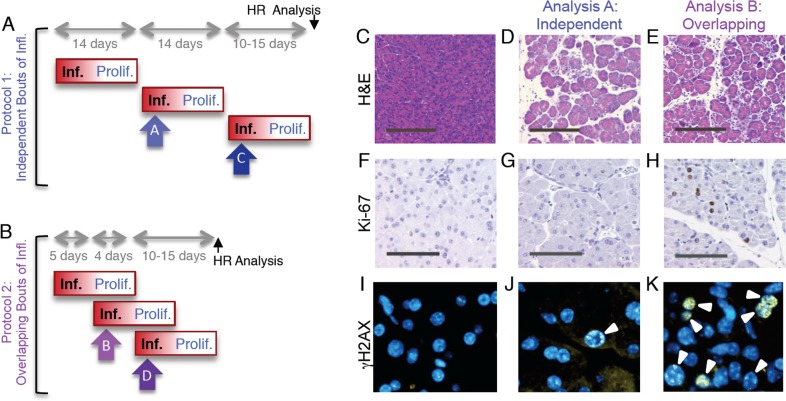
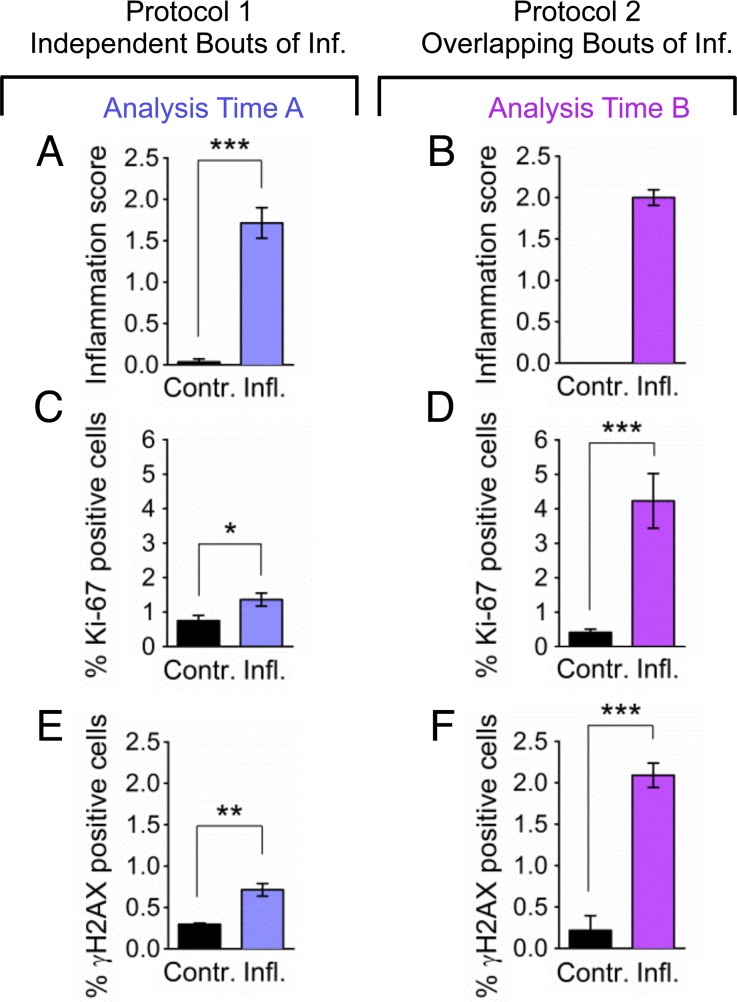
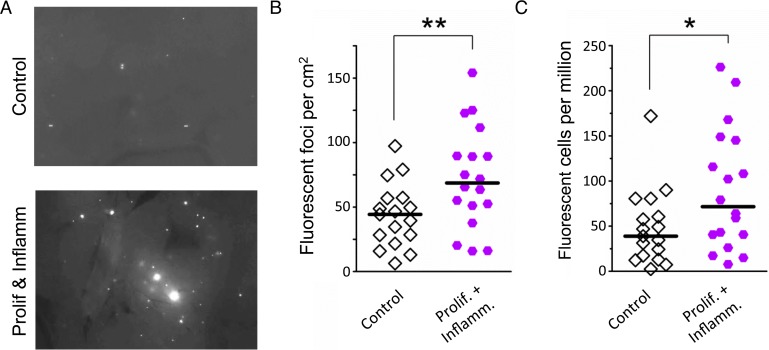
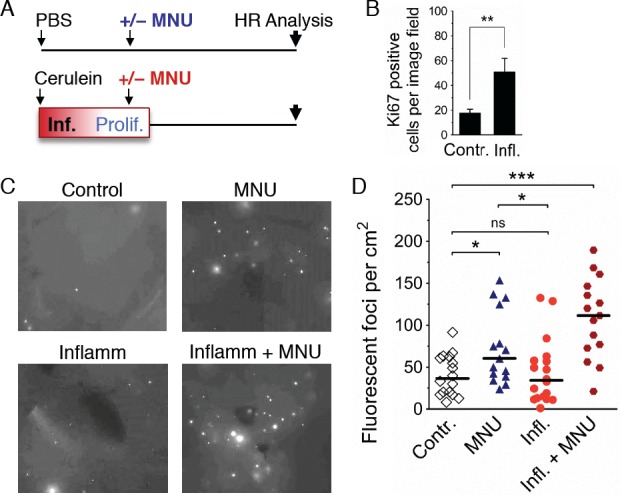
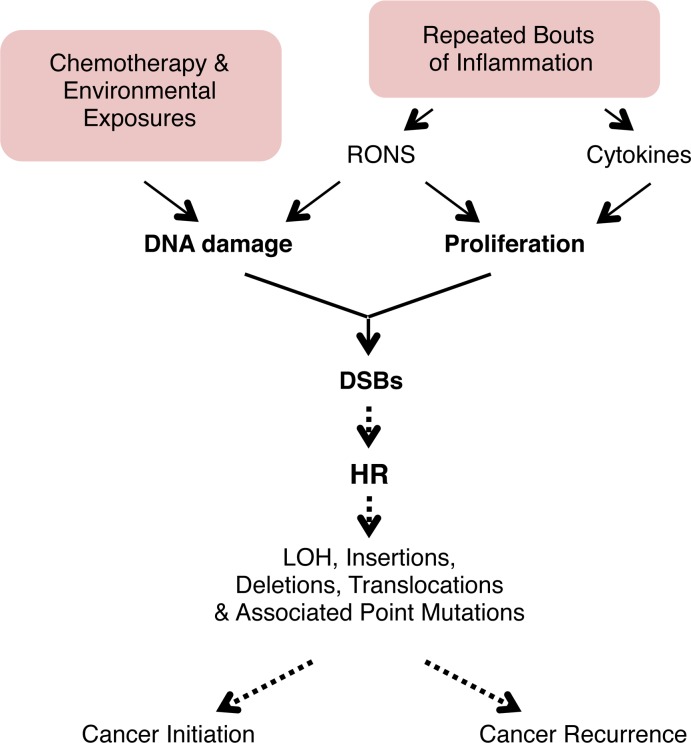
Mutations are a critical driver of cancer initiation. While extensive studies have focused on exposure-induced mutations, few studies have explored the importance of tissue physiology as a modulator of mutation susceptibility in vivo. Of particular interest is inflammation, a known cancer risk factor relevant to chronic inflammatory diseases and pathogen-induced inflammation. Here, we used the fluorescent yellow direct repeat (FYDR) mice that harbor a reporter to detect misalignments during homologous recombination (HR), an important class of mutations. FYDR mice were exposed to cerulein, a potent inducer of pancreatic inflammation. We show that inflammation induces DSBs (γH2AX foci) and that several days later there is an increase in cell proliferation. While isolated bouts of inflammation did not induce HR, overlap between inflammation-induced DNA damage and inflammation-induced cell proliferation induced HR significantly. To study exogenously-induced DNA damage, animals were exposed to methylnitrosourea, a model alkylating agent that creates DNA lesions relevant to both environmental exposures and cancer chemotherapy. We found that exposure to alkylation damage induces HR, and importantly, that inflammation-induced cell proliferation and alkylation induce HR in a synergistic fashion. Taken together, these results show that, during an acute bout of inflammation, there is a kinetic barrier separating DNA damage from cell proliferation that protects against mutations, and that inflammation-induced cell proliferation greatly potentiates exposure-induced mutations. These studies demonstrate a fundamental mechanism by which inflammation can act synergistically with DNA damage to induce mutations that drive cancer and cancer recurrence.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Friedberg EC, Walker GC, Siede W, Wood RD, Schultz R, and Ellenberger T. (2006). DNA repair and mutagenesis. ASM Press, Washington, DC, USA.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
