

Membrane recognition and dynamics of the RNA degradosome
- PMID: 25647427
- PMCID: PMC4372235
- DOI: 10.1371/journal.pgen.1004961
Membrane recognition and dynamics of the RNA degradosome
Abstract
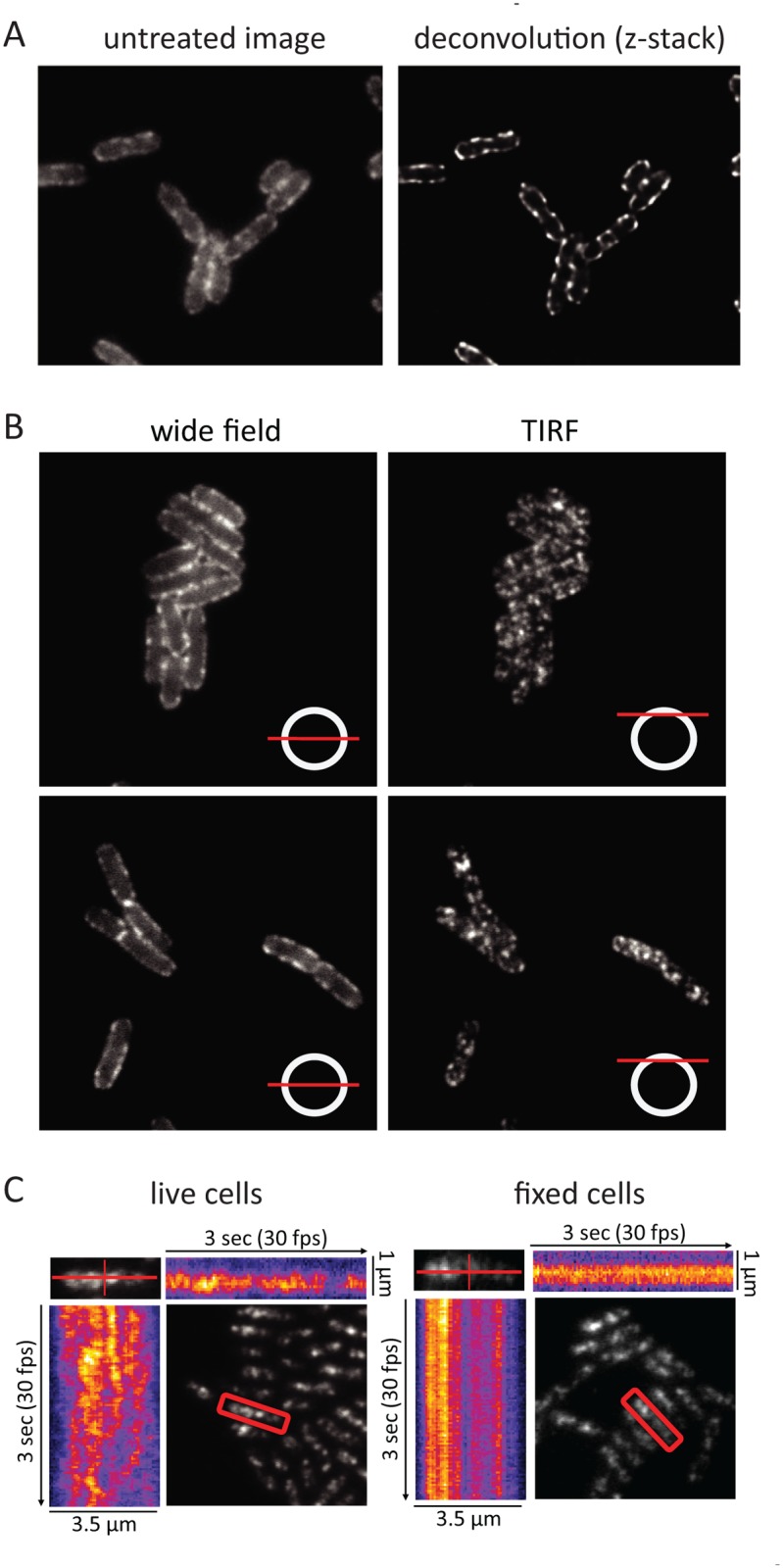
RNase E, which is the central component of the multienzyme RNA degradosome, serves as a scaffold for interaction with other enzymes involved in mRNA degradation including the DEAD-box RNA helicase RhlB. Epifluorescence microscopy under live cell conditions shows that RNase E and RhlB are membrane associated, but neither protein forms cytoskeletal-like structures as reported earlier by Taghbalout and Rothfield. We show that association of RhlB with the membrane depends on a direct protein interaction with RNase E, which is anchored to the inner cytoplasmic membrane through an MTS (Membrane Targeting Sequence). Molecular dynamics simulations show that the MTS interacts with the phospholipid bilayer by forming a stabilized amphipathic α-helix with the helical axis oriented parallel to the plane of the bilayer and hydrophobic side chains buried deep in the acyl core of the membrane. Based on the molecular dynamics simulations, we propose that the MTS freely diffuses in the membrane by a novel mechanism in which a large number of weak contacts are rapidly broken and reformed. TIRFm (Total Internal Reflection microscopy) shows that RNase E in live cells rapidly diffuses over the entire inner membrane forming short-lived foci. Diffusion could be part of a scanning mechanism facilitating substrate recognition and cooperativity. Remarkably, RNase E foci disappear and the rate of RNase E diffusion increases with rifampicin treatment. Control experiments show that the effect of rifampicin is specific to RNase E and that the effect is not a secondary consequence of the shut off of E. coli transcription. We therefore interpret the effect of rifampicin as being due to the depletion of RNA substrates for degradation. We propose a model in which formation of foci and constraints on diffusion arise from the transient clustering of RNase E into cooperative degradation bodies.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
