

Host-based Prophylaxis Successfully Targets Liver Stage Malaria Parasites
- PMID: 25648263
- PMCID: PMC4427874
- DOI: 10.1038/mt.2015.18
Host-based Prophylaxis Successfully Targets Liver Stage Malaria Parasites
Abstract
Eliminating malaria parasites during the asymptomatic but obligate liver stages (LSs) of infection would stop disease and subsequent transmission. Unfortunately, only a single licensed drug that targets all LSs, Primaquine, is available. Targeting host proteins might significantly expand the repertoire of prophylactic drugs against malaria. Here, we demonstrate that both Bcl-2 inhibitors and P53 agonists dramatically reduce LS burden in a mouse malaria model in vitro and in vivo by altering the activity of key hepatocyte factors on which the parasite relies. Bcl-2 inhibitors act primarily by inducing apoptosis in infected hepatocytes, whereas P53 agonists eliminate parasites in an apoptosis-independent fashion. In combination, Bcl-2 inhibitors and P53 agonists act synergistically to delay, and in some cases completely prevent, the onset of blood stage disease. Both families of drugs are highly effective at doses that do not cause substantial hepatocyte cell death in vitro or liver damage in vivo. P53 agonists and Bcl-2 inhibitors were also effective when administered to humanized mice infected with Plasmodium falciparum. Our data demonstrate that host-based prophylaxis could be developed into an effective intervention strategy that eliminates LS parasites before the onset of clinical disease and thus opens a new avenue to prevent malaria.
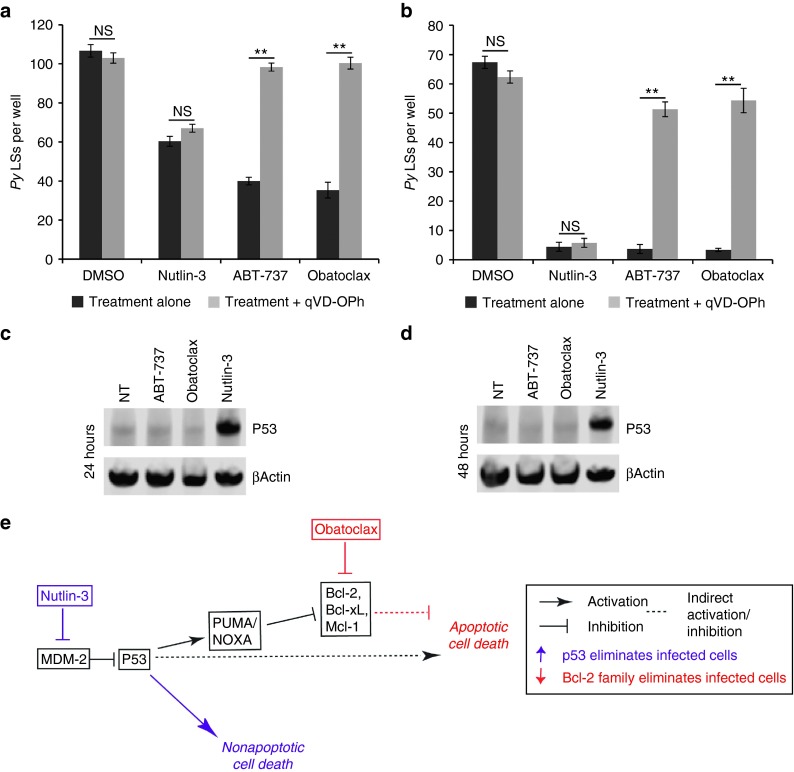
Figures




Similar articles
-
Assessing drug efficacy against Plasmodium falciparum liver stages in vivo.JCI Insight. 2018 Jan 11;3(1):e92587. doi: 10.1172/jci.insight.92587. eCollection 2018 Jan 11. JCI Insight. 2018. PMID: 29321371 Free PMC article.
-
Imaging of Plasmodium liver stages to drive next-generation antimalarial drug discovery.Science. 2011 Dec 9;334(6061):1372-7. doi: 10.1126/science.1211936. Epub 2011 Nov 17. Science. 2011. PMID: 22096101 Free PMC article.
-
Suppression of host p53 is critical for Plasmodium liver-stage infection.Cell Rep. 2013 Mar 28;3(3):630-7. doi: 10.1016/j.celrep.2013.02.010. Epub 2013 Mar 7. Cell Rep. 2013. PMID: 23478020 Free PMC article.
-
Progressing the global antimalarial portfolio: finding drugs which target multiple Plasmodium life stages.Parasitology. 2014 Jan;141(1):66-76. doi: 10.1017/S0031182013000747. Epub 2013 Jun 10. Parasitology. 2014. PMID: 23746048 Review.
-
That was then but this is now: malaria research in the time of an eradication agenda.Science. 2010 May 14;328(5980):862-6. doi: 10.1126/science.1184785. Science. 2010. PMID: 20466924 Review.
Cited by
-
The Promise of Systems Biology Approaches for Revealing Host Pathogen Interactions in Malaria.Front Microbiol. 2017 Nov 16;8:2183. doi: 10.3389/fmicb.2017.02183. eCollection 2017. Front Microbiol. 2017. PMID: 29201016 Free PMC article. Review.
-
HGF Secreted by Activated Kupffer Cells Induces Apoptosis of Plasmodium-Infected Hepatocytes.Front Immunol. 2017 Feb 6;8:90. doi: 10.3389/fimmu.2017.00090. eCollection 2017. Front Immunol. 2017. PMID: 28220125 Free PMC article.
-
Selection and refinement: the malaria parasite's infection and exploitation of host hepatocytes.Curr Opin Microbiol. 2015 Aug;26:71-8. doi: 10.1016/j.mib.2015.05.013. Epub 2015 Jun 21. Curr Opin Microbiol. 2015. PMID: 26102161 Free PMC article. Review.
-
Discovery of Druggable Host Factors Critical to Plasmodium Liver-Stage Infection.Cell Chem Biol. 2019 Sep 19;26(9):1253-1262.e5. doi: 10.1016/j.chembiol.2019.05.011. Epub 2019 Jun 27. Cell Chem Biol. 2019. PMID: 31257182 Free PMC article.
-
Elucidating Spatially-Resolved Changes in Host Signaling During Plasmodium Liver-Stage Infection.Front Cell Infect Microbiol. 2022 Jan 17;11:804186. doi: 10.3389/fcimb.2021.804186. eCollection 2021. Front Cell Infect Microbiol. 2022. PMID: 35111697 Free PMC article.
References
-
- Kappe SH, Vaughan AM, Boddey JA, Cowman AF. That was then but this is now: malaria research in the time of an eradication agenda. Science. 2010;328:862–866. - PubMed
-
- Muregi FW. Antimalarial drugs and their useful therapeutic lives: rational drug design lessons from pleiotropic action of quinolines and artemisinins. Curr Drug Discov Technol. 2010;7:280–316. - PubMed
-
- Arrow KJ, Panosian C, Gelband H.2004Arrow KJ, Panosian C, Gelband H.eds). Saving Lives, Buying Time: Economics of Malaria Drugs in an Age of Resistance.Washington, DC: National Academies Press. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
