

α-2,3-sialyltransferase expression level impacts the kinetics of lipooligosaccharide sialylation, complement resistance, and the ability of Neisseria gonorrhoeae to colonize the murine genital tract
- PMID: 25650401
- PMCID: PMC4324315
- DOI: 10.1128/mBio.02465-14
α-2,3-sialyltransferase expression level impacts the kinetics of lipooligosaccharide sialylation, complement resistance, and the ability of Neisseria gonorrhoeae to colonize the murine genital tract
Abstract
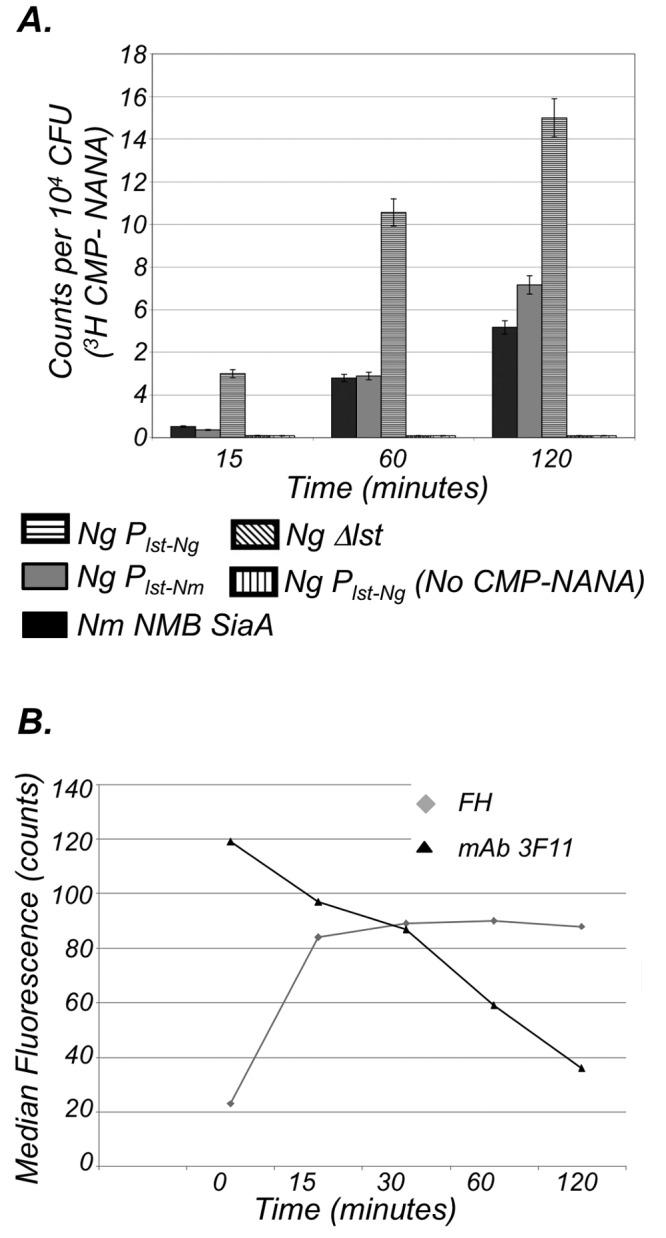
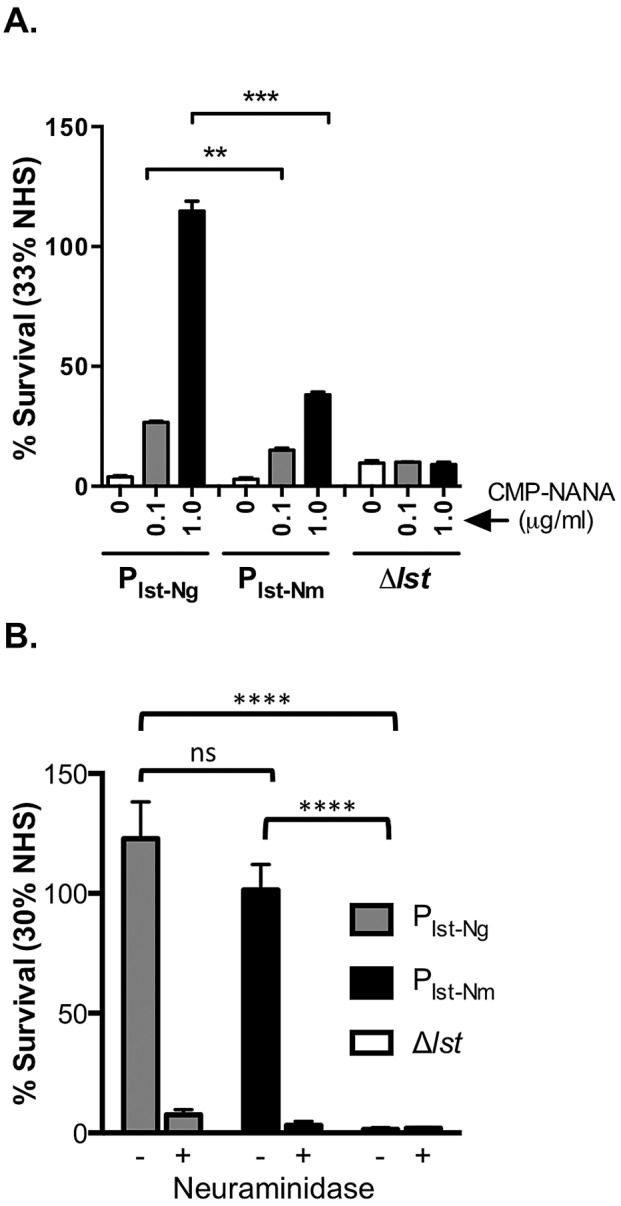
Neisseria meningitidis and Neisseria gonorrhoeae modify the terminal lacto-N-neotetraose moiety of their lipooligosaccharide (LOS) with sialic acid. N. gonorrhoeae LOS sialylation blocks killing by complement, which is mediated at least in part by enhanced binding of the complement inhibitor factor H (FH). The role of LOS sialylation in resistance of N. meningitidis to serum killing is less well defined. Sialylation in each species is catalyzed by the enzyme LOS α-2,3-sialyltransferase (Lst). Previous studies have shown increased Lst activity in N. gonorrhoeae compared to N. meningitidis due to an ~5-fold increase in lst transcription. Using isogenic N. gonorrhoeae strains engineered to express gonococcal lst from either the N. gonorrhoeae or N. meningitidis lst promoter, we show that decreased expression of lst (driven by the N. meningitidis promoter) reduced LOS sialylation as determined by less incorporation of tritium-labeled cytidine monophospho-N-acetylneuraminic acid (CMP-NANA; the donor molecule for sialic acid). Diminished LOS sialylation resulted in reduced rates of FH binding and increased pathway activation compared to N. gonorrhoeae promoter-driven lst expression. The N. meningitidis lst promoter generated sufficient Lst to sialylate N. gonorrhoeae LOS in vivo, and the level of sialylation after 24 h in the mouse genital tract was sufficient to mediate resistance to human serum ex vivo. Despite demonstrable LOS sialylation in vivo, gonococci harboring the N. meningitidis lst promoter were outcompeted by those with the N. gonorrhoeae lst promoter during coinfection of the vaginal tract of estradiol-treated mice. These data highlight the importance of high lst expression levels for gonococcal pathogenesis.
Importance: Neisseria gonorrhoeae has become resistant to nearly every therapeutic antibiotic used and is listed as an "urgent threat" by the Centers for Disease Control and Prevention. Novel therapies are needed to combat drug-resistant N. gonorrhoeae. Gonococci express an α-2,3-sialyltransferase (Lst) that can scavenge sialic acid from the host and use it to modify lipooligosaccharide (LOS). Sialylation of gonococcal LOS converts serum-sensitive strains to serum resistance, decreases antibody binding, and combats killing by neutrophils and antimicrobial peptides. Mutant N. gonorrhoeae that lack Lst (cannot sialylate LOS) are attenuated in a mouse model. Lst expression levels differ among N. gonorrhoeae strains, and N. gonorrhoeae typically expresses more Lst than Neisseria meningitidis. Here we examined the significance of differential lst expression levels and determined that the level of LOS sialylation is critical to the ability of N. gonorrhoeae to combat the immune system and survive in an animal model. LOS sialylation may be an ideal target for novel therapies.
Copyright © 2015 Lewis et al.
Figures


References
-
- Joiner KA, Warren KA, Brown EJ, Swanson J, Frank MM. 1983. Studies on the mechanism of bacterial resistance to complement-mediated killing. IV. C5b-9 forms high molecular weight complexes with bacterial outer membrane constituents on serum-resistant but not on serum-sensitive Neisseria gonorrhoeae. J Immunol 131:1443–1451. - PubMed
-
- Giuntini S, Reason DC, Granoff DM. 2012. Combined roles of human IgG subclass, alternative complement pathway activation, and epitope density in the bactericidal activity of antibodies to meningococcal factor H binding protein. Infect Immun 80:187–194. doi: 10.1128/IAI.05956-11. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
