

MBTPS1/SKI-1/S1P proprotein convertase is required for ECM signaling and axial elongation during somitogenesis and vertebral development†
- PMID: 25652402
- PMCID: PMC4406298
- DOI: 10.1093/hmg/ddv050
MBTPS1/SKI-1/S1P proprotein convertase is required for ECM signaling and axial elongation during somitogenesis and vertebral development†
Abstract
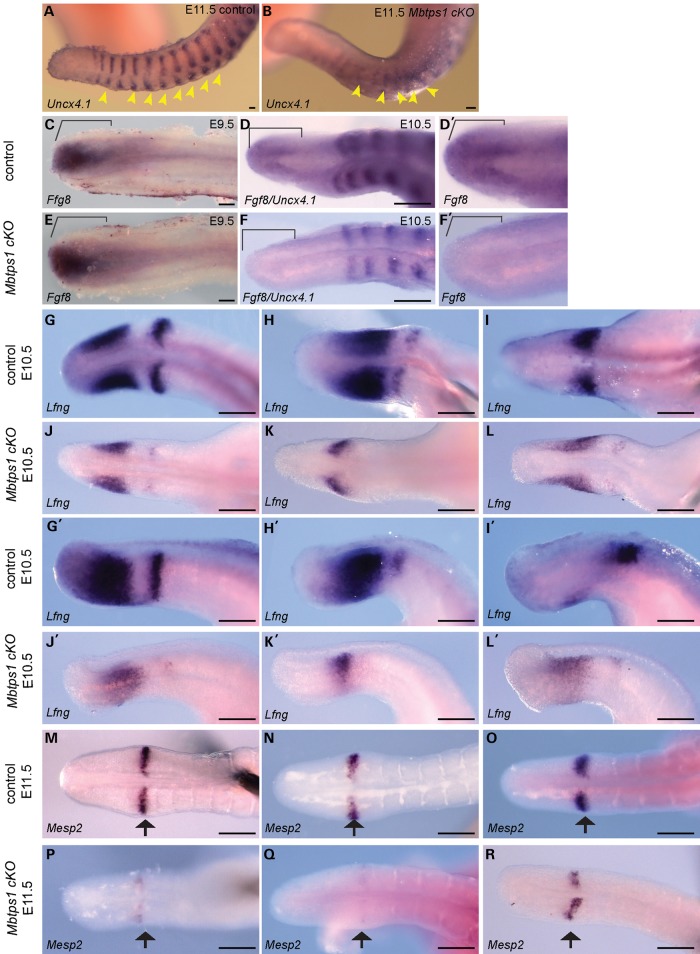
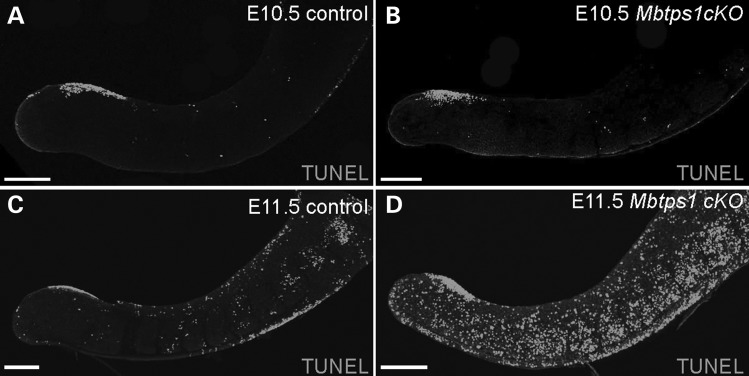
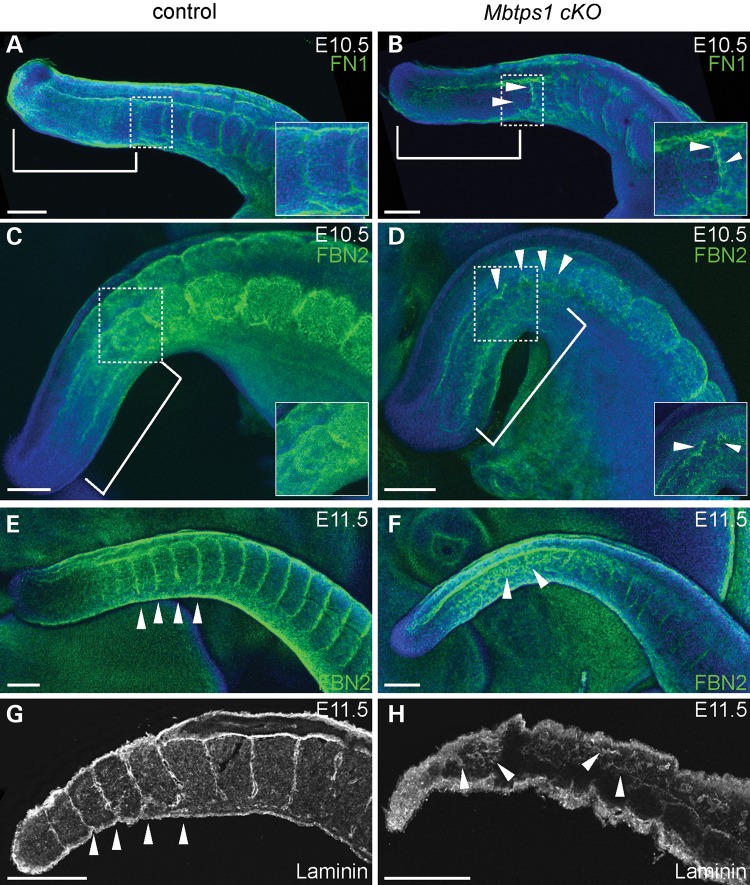
Caudal regression syndrome (sacral agenesis), which impairs development of the caudal region of the body, occurs with a frequency of about 2 live births per 100 000 newborns although this incidence rises to 1 in 350 infants born to mothers with gestational diabetes. The lower back and limbs can be affected as well as the genitourinary and gastrointestinal tracts. The axial skeleton is formed during embryogenesis through the process of somitogenesis in which the paraxial mesoderm periodically segments into bilateral tissue blocks, called somites. Somites are the precursors of vertebrae and associated muscle, tendons and dorsal dermis. Vertebral anomalies in caudal regression syndrome may arise through perturbation of somitogenesis or, alternatively, could result from defective bone formation and patterning. We discovered that MBTPS1/SKI-1/S1P, which proteolytically activates a class of transmembrane transcription factors, plays a critical role in somitogenesis and the pathogenesis of lumbar/sacral vertebral anomalies. Conditional deletion of Mbtps1 yields a viable mouse with misshapen, fused and reduced number of lumbar and sacral vertebrae, under-developed hind limb bones and a kinky, shortened tail. We show that Mbtps1 is required to (i) maintain the Fgf8 'wavefront' in the presomitic mesoderm that underpins axial elongation, (ii) sustain the Lfng oscillatory 'clock' activity that governs the periodicity of somite formation and (iii) preserve the composition and character of the somitic extracellular matrix containing fibronectin, fibrillin2 and laminin. Based on this spinal phenotype and known functions of MBTPS1, we reason that loss-of-function mutations in Mbtps1 may cause the etiology of caudal regression syndrome.
© The Author 2015. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
References
-
- Boulas M.M. (2009) Recognition of caudal regression syndrome. Adv. Neonatal. Care 9, 61–69 quiz 70-61. - PubMed
-
- Thottungal A.D., Charles A.K., Dickinson J.E., Bower C. (2010) Caudal dysgenesis and sirenomelia-single centre experience suggests common pathogenic basis. Am. J. Med. Genet. A, 152A, 2578–2587. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
