

X-ray structure of the pestivirus NS3 helicase and its conformation in solution
- PMID: 25653438
- PMCID: PMC4442355
- DOI: 10.1128/JVI.03165-14
X-ray structure of the pestivirus NS3 helicase and its conformation in solution
Abstract
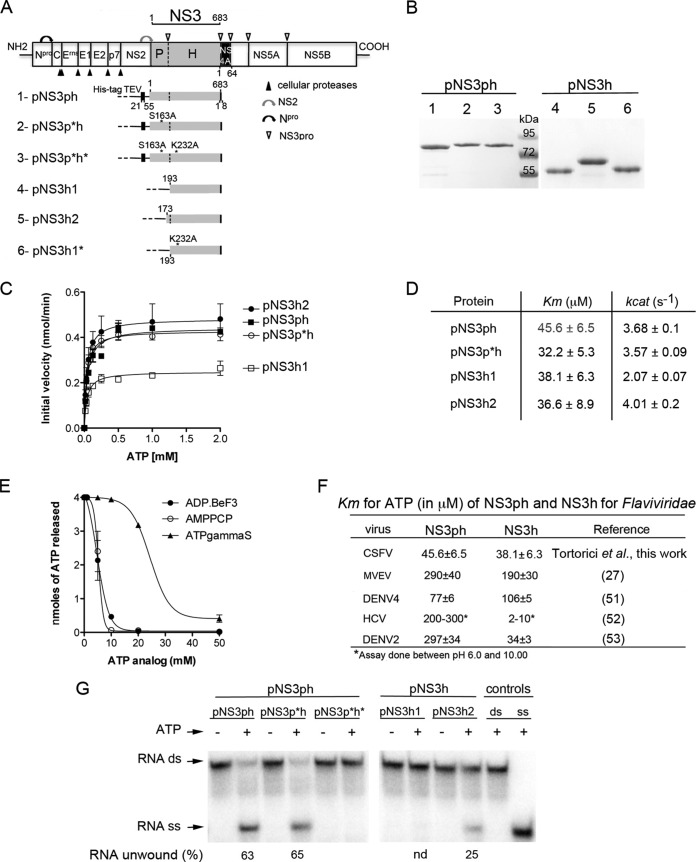
Pestiviruses form a genus in the Flaviviridae family of small enveloped viruses with a positive-sense single-stranded RNA genome. Viral replication in this family requires the activity of a superfamily 2 RNA helicase contained in the C-terminal domain of nonstructural protein 3 (NS3). NS3 features two conserved RecA-like domains (D1 and D2) with ATPase activity, plus a third domain (D3) that is important for unwinding nucleic acid duplexes. We report here the X-ray structure of the pestivirus NS3 helicase domain (pNS3h) at a 2.5-Å resolution. The structure deviates significantly from that of NS3 of other genera in the Flaviviridae family in D3, as it contains two important insertions that result in a narrower nucleic acid binding groove. We also show that mutations in pNS3h that rescue viruses from which the core protein is deleted map to D3, suggesting that this domain may be involved in interactions that facilitate particle assembly. Finally, structural comparisons of the enzyme in different crystalline environments, together with the findings of small-angle X-ray-scattering studies in solution, show that D2 is mobile with respect to the rest of the enzyme, oscillating between closed and open conformations. Binding of a nonhydrolyzable ATP analog locks pNS3h in a conformation that is more compact than the closest apo-form in our crystals. Together, our results provide new insight and bring up new questions about pNS3h function during pestivirus replication.
Importance: Although pestivirus infections impose an important toll on the livestock industry worldwide, little information is available about the nonstructural proteins essential for viral replication, such as the NS3 helicase. We provide here a comparative structural and functional analysis of pNS3h with respect to its orthologs in other viruses of the same family, the flaviviruses and hepatitis C virus. Our studies reveal differences in the nucleic acid binding groove that could have implications for understanding the unwinding specificity of pNS3h, which is active only on RNA duplexes. We also show that pNS3h has a highly dynamic behavior--a characteristic probably shared with NS3 helicases from all Flaviviridae members--that could be targeted for drug design by using recent algorithms to specifically block molecular motion. Compounds that lock the enzyme in a single conformation or limit its dynamic range of conformations are indeed likely to block its helicase function.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures





Similar articles
-
A positive-strand RNA virus uses alternative protein-protein interactions within a viral protease/cofactor complex to switch between RNA replication and virion morphogenesis.PLoS Pathog. 2017 Feb 2;13(2):e1006134. doi: 10.1371/journal.ppat.1006134. eCollection 2017 Feb. PLoS Pathog. 2017. PMID: 28151973 Free PMC article.
-
Uncoupling of Protease trans-Cleavage and Helicase Activities in Pestivirus NS3.J Virol. 2017 Oct 13;91(21):e01094-17. doi: 10.1128/JVI.01094-17. Print 2017 Nov 1. J Virol. 2017. PMID: 28835495 Free PMC article.
-
Nucleoside triphosphatase and RNA helicase activities associated with GB virus B nonstructural protein 3.Virology. 1999 Sep 1;261(2):216-26. doi: 10.1006/viro.1999.9871. Virology. 1999. PMID: 10497107
-
Structure and flexibility of non-structural proteins 3 and -5 of Dengue- and Zika viruses in solution.Prog Biophys Mol Biol. 2019 May;143:67-77. doi: 10.1016/j.pbiomolbio.2018.08.008. Epub 2018 Aug 29. Prog Biophys Mol Biol. 2019. PMID: 30171868 Review.
-
Functional interplay among the flavivirus NS3 protease, helicase, and cofactors.Virol Sin. 2014 Apr;29(2):74-85. doi: 10.1007/s12250-014-3438-6. Epub 2014 Mar 26. Virol Sin. 2014. PMID: 24691778 Free PMC article. Review.
Cited by
-
Characterization of the Determinants of NS2-3-Independent Virion Morphogenesis of Pestiviruses.J Virol. 2015 Nov;89(22):11668-80. doi: 10.1128/JVI.01646-15. Epub 2015 Sep 9. J Virol. 2015. PMID: 26355097 Free PMC article.
-
A positively charged surface patch on the pestivirus NS3 protease module plays an important role in modulating NS3 helicase activity and virus production.Arch Virol. 2021 Jun;166(6):1633-1642. doi: 10.1007/s00705-021-05055-5. Epub 2021 Mar 31. Arch Virol. 2021. PMID: 33787991
-
Structural Virology: The Key Determinants in Development of Antiviral Therapeutics.Viruses. 2025 Mar 14;17(3):417. doi: 10.3390/v17030417. Viruses. 2025. PMID: 40143346 Free PMC article. Review.
-
NSUN2-mediated HCV RNA m5C Methylation Facilitates Viral RNA Stability and Replication.Genomics Proteomics Bioinformatics. 2025 May 10;23(1):qzaf008. doi: 10.1093/gpbjnl/qzaf008. Genomics Proteomics Bioinformatics. 2025. PMID: 39957240 Free PMC article.
-
A positive-strand RNA virus uses alternative protein-protein interactions within a viral protease/cofactor complex to switch between RNA replication and virion morphogenesis.PLoS Pathog. 2017 Feb 2;13(2):e1006134. doi: 10.1371/journal.ppat.1006134. eCollection 2017 Feb. PLoS Pathog. 2017. PMID: 28151973 Free PMC article.
References
-
- Lindenbach BM, Murray CL,Thiel HJ, Rice CM. 2013. Flaviviridae, p 712–746. Knipe DM, Howley PM, Cohen JI, Griffin DE, Lamb RA, Martin MA, Racaniello VR, Roizman B (ed), Fields virology, 6th ed Lippincott Williams & Wilkins, Philadelphia, PA.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
