

Emerging critical roles of Fe-S clusters in DNA replication and repair
- PMID: 25655665
- PMCID: PMC4576882
- DOI: 10.1016/j.bbamcr.2015.01.018
Emerging critical roles of Fe-S clusters in DNA replication and repair
Abstract
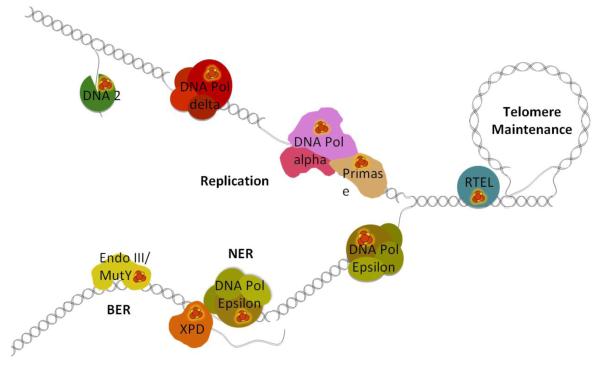
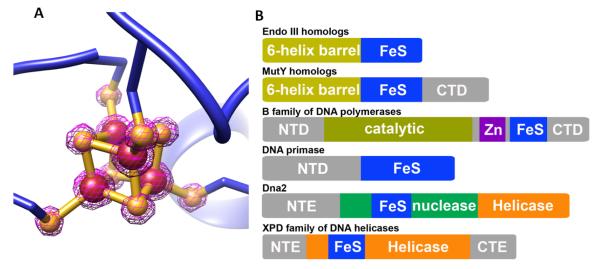
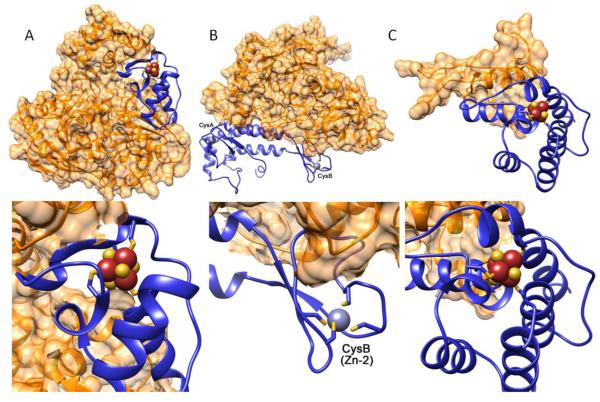
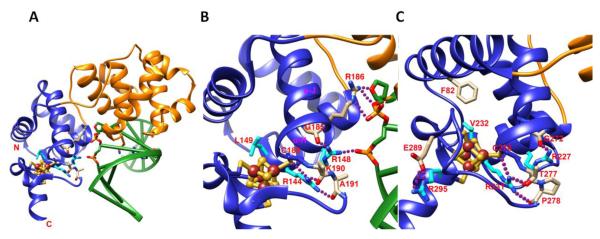
Fe-S clusters are partners in the origin of life that predate cells, acetyl-CoA metabolism, DNA, and the RNA world. The double helix solved the mystery of DNA replication by base pairing for accurate copying. Yet, for genome stability necessary to life, the double helix has equally important implications for damage repair. Here we examine striking advances that uncover Fe-S cluster roles both in copying the genetic sequence by DNA polymerases and in crucial repair processes for genome maintenance, as mutational defects cause cancer and degenerative disease. Moreover, we examine an exciting, controversial role for Fe-S clusters in a third element required for life - the long-range coordination and regulation of replication and repair events. By their ability to delocalize electrons over both Fe and S centers, Fe-S clusters have unbeatable features for protein conformational control and charge transfer via double-stranded DNA that may fundamentally transform our understanding of life, replication, and repair. This article is part of a Special Issue entitled: Fe/S proteins: Analysis, structure, function, biogenesis and diseases.
Keywords: Cancer and degenerative disease; DNA charge transfer communication; DNA repair; DNA replication; Fe–S cluster; Genome integrity.
Copyright © 2015 Elsevier B.V. All rights reserved.
Figures
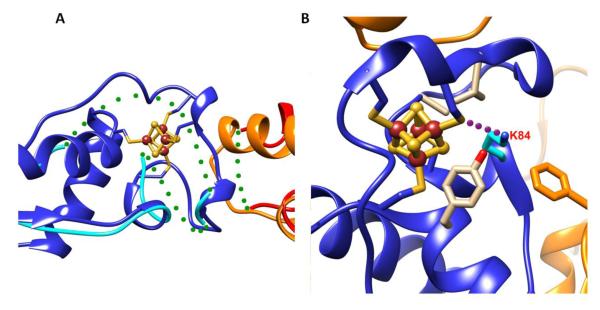
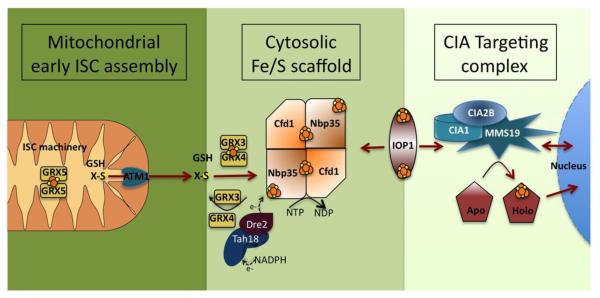
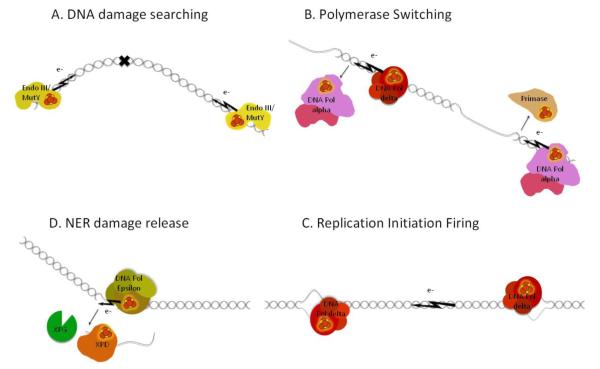
References
-
- Fontecave M. Iron-sulfur clusters: ever-expanding roles. Nat Chem Biol. 2006;2:171–174. doi:10.1038/nchembio0406-171. - PubMed
-
- Imlay JA. Iron-sulphur clusters and the problem with oxygen. Mol Microbiol. 2006;59:1073–1082. doi:10.1111/j.1365-2958.2006.05028.x. - PubMed
-
- Russell MJ, Martin W. The rocky roots of the acetyl-CoA pathway. Trends Biochem Sci. 2004;29:358–363. doi:10.1016/j.tibs.2004.05.007. - PubMed
-
- Imlay JA. Pathways of oxidative damage. Annu. Rev. Microbiol. 2003;57:395–418. doi:10.1146/annurev.micro.57.030502.090938. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
