

Dorsoventral differences in Kv7/M-current and its impact on resonance, temporal summation and excitability in rat hippocampal pyramidal cells
- PMID: 25656084
- PMCID: PMC4386960
- DOI: 10.1113/jphysiol.2014.280826
Dorsoventral differences in Kv7/M-current and its impact on resonance, temporal summation and excitability in rat hippocampal pyramidal cells
Abstract
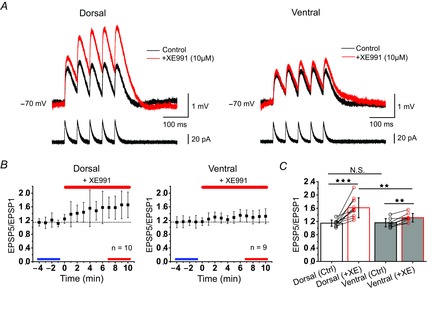
In rodent hippocampi, the connections, gene expression and functions differ along the dorsoventral (D-V) axis. CA1 pyramidal cells show increasing excitability along the D-V axis, although the underlying mechanism is not known. In the present study, we investigated how the M-current (IM ), caused by Kv7/M (KCNQ) potassium channels, and known to often control neuronal excitability, contributes to D-V differences in intrinsic properties of CA1 pyramidal cells. Using whole-cell patch clamp recordings and the selective Kv7/M blocker 10,10-bis(4-pyridinylmethyl)-9(10H)-anthracenone dihydrochloride (XE991) in hippocampal slices from 3- to 4-week-old rats, we found that: (i) IM had a stronger impact on subthreshold electrical properties in dorsal than ventral CA1 pyramidal cells, including input resistance, temporal summation of artificial synaptic potentials, and M-resonance; (ii) IM activated at more negative potentials (left-shifted) and had larger peak amplitude in the dorsal than ventral CA1; and (iii) the initial spike threshold (during ramp depolarizations) was elevated, and the medium after-hyperpolarization and spike frequency adaptation were increased (i.e. excitability was lower) in the dorsal rather than ventral CA1. These differences were abolished or reduced by application of XE991, indicating that they were caused by IM . Thus, it appears that IM has stronger effects in dorsal than in ventral rat CA1 pyramidal cells because of a larger maximal M-conductance and left-shifted activation curve in the dorsal cells. These mechanisms may contribute to D-V differences in the rate and phase coding of position by CA1 place cells, and may also enhance epileptiform activity in ventral CA1.
© 2014 The Authors. The Journal of Physiology © 2014 The Physiological Society.
Figures










Comment in
-
Computational diversity in the hippocampus: a matter of components.J Physiol. 2015 Apr 1;593(7):1525-6. doi: 10.1113/jphysiol.2014.288456. J Physiol. 2015. PMID: 25828642 Free PMC article. No abstract available.
References
-
- Alle H, Ostroumov K, Geiger J. Storm JF. M-current, persistent Na+ current, and subthreshold resonance recorded in mossy fiber boutons. 2009. (MFBs) in rat hippocampus. Program No. 42.2/D10. 2009 Neuroscience Meeting Planner. Society for Neuroscience, Chicago, IL, USA. Online. Ref Type: Abstract.
-
- Bannerman DM, Rawlins JNP, McHugh SB, Deacon RMJ, Yee BK, Bast T, Zhang WN, Pothuizen HHJ. Feldon J. Regional dissociations within the hippocampus – memory and anxiety. Neurosci Biobehav Rev. 2004;28:273–283. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous