

Review
doi: 10.1038/nri3804.
Epub 2015 Feb 6.
Dynamics of B cells in germinal centres
Affiliations
- PMID: 25656706
- PMCID: PMC4399774
- DOI: 10.1038/nri3804
Item in Clipboard
Review
Dynamics of B cells in germinal centres
Nat Rev Immunol.
2015 Mar.
Abstract
Humoral immunity depends on the germinal centre (GC) reaction during which somatically mutated high-affinity memory B cells and plasma cells are generated. Recent studies have uncovered crucial cues that are required for the formation and the maintenance of GCs and for the selection of high-affinity antibody mutants. In addition, it is now clear that these events are promoted by the dynamic movements of cells within and between GCs. These findings have resolved the complexities of the GC reaction in greater detail than ever before. This Review focuses on these recent advances and discusses their implications for the establishment of humoral immunity.
Figures
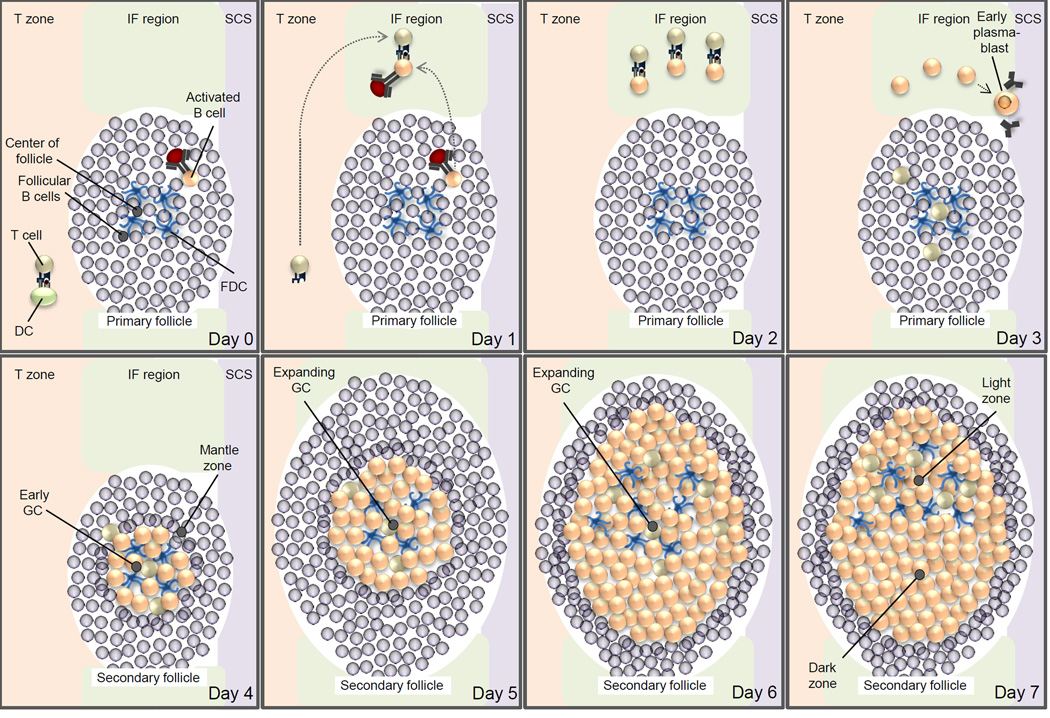
This schematic figure is primarily based on two recent studies that determined the movements of antigen-specific B cells and T cells in lymph nodes during the immune response to a model antigen,. On day 0, B cells and T cells are activated by recognition of their cognate antigen in the primary follicle and T cell zone, respectively. On day 1, the activated B cells and T cells migrate to the interfollicular region and start to interact. On day 2, B cells and T cells form long-lived interactions, resulting in the full activation of B cells. T cells acquire the characteristic T follicular helper cell (TFH cell) phenotype. On day 3, TFH cells migrate from the interfollicular region into the follicle. Some antigen-activated B cells differentiate into antibody-secreting cells or early plasmablasts that move to a region adjacent to the subcapsular sinus (SCS). On day 4, B cells migrate from the interfollicular region into the centre of the follicle — which is characterized by a network of follicular dendritic cells (FDCs) — begin to proliferate and, as a result, push the resident follicular B cells aside to form the early germinal centre (GC), which consists of B cell blasts surrounded by the mantle zone. This structure is also referred to as the secondary follicle, thus distinguishing it from the ‘GC-less’ primary follicle. On days 5–6, the GC rapidly expands as a result of the fast proliferation of the B cell blasts. On day 7, dark zones and light zones form, which results in the establishment of the mature GC. The dark zone mainly consists of densely packed B cell blasts, whereas the light zone contains TFH cells and FDCs. CD40L, CD40 ligand; DC, dendritic cell; TCR, T cell receptor.
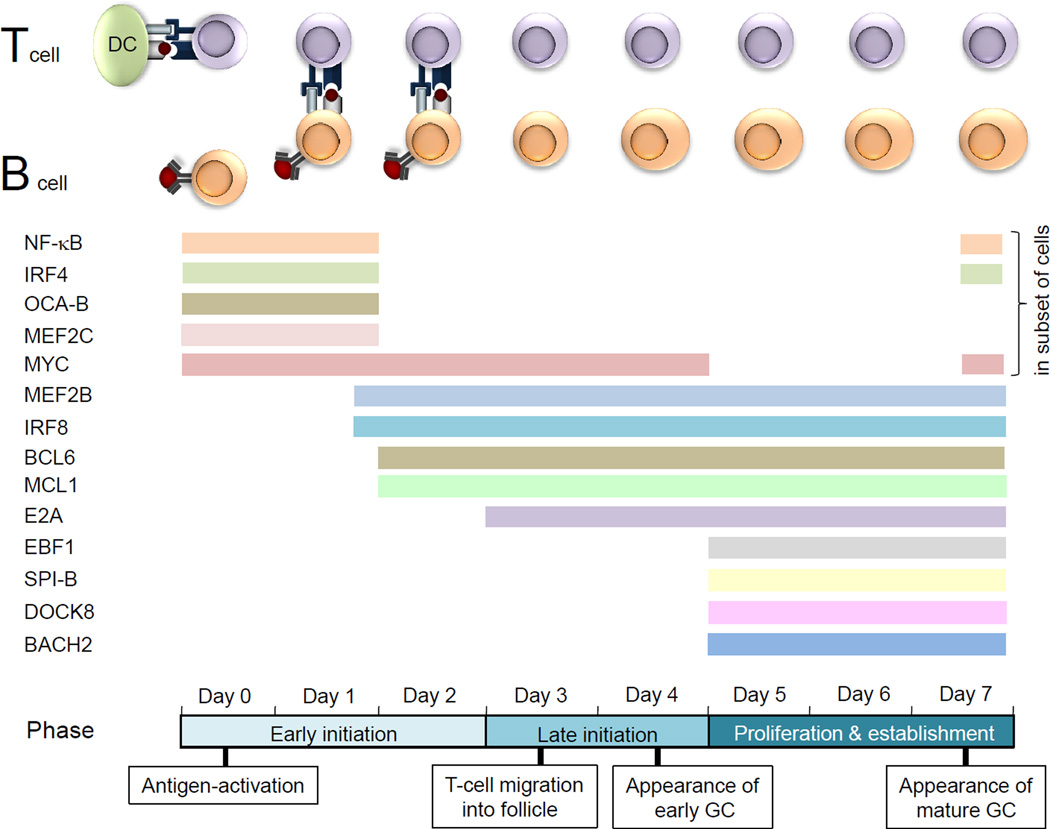
Published findings are consistent with the existence of three separate phases during the first 7 days of the germinal centre (GC) reaction. An early initiation phase (days 0–2) encompasses the activation of the B cells and T cells by their cognate antigen and the long-lived interactions of B cells and T cells in the interfollicular region. A late initiation phase (days 3–4) is characterized by the migration of the T cells first and 1 day later the migration of the B cells out of the interfollicular region and the appearance of the early GC. A proliferation and establishment phase (days 5–7) occurs in which the GC grows and eventually forms the characteristic dark and light zone patterning of the mature GC. Studies of the GC reaction following the deletion of molecules in antigen-activated or GC B cells have revealed a functional requirement for: nuclear factor-κB (NF-κB); interferon-regulatory factor 4 (IRF4); OCA-B; myocyte-specific enhancer factor 2C (MEF2C); MYC; B cell lymphoma 6 (BCL-6); myeloid cell leukaemia 1 (MCL1); E2A; early B cell factor 1 (EBF1); SPIB; and dedicator of cytokinesis protein 8 (DOCK8). A functional role has also been suggested for several other factors: MEF2B, on the basis of its expression pattern in antigen-activated B cells in vivo; IRF8, on the basis of its high expression in GC B cells and its role in the induction of BCL-6 expression; and BACH2 (BTB and CNC homologue 2), on the basis of its expression in established GC B cells (earlier time points have not been analysed). The exact time point at which BACH2 deficiency affects the formation of GCs in not known. As BACH2 cooperates with BCL-6 in the suppression of genes in GC B cells, we propose that BACH2 is required in the proliferation and establishment phase, but note that it could also be required in an earlier phase. CD40L, CD40 ligand; DC, dendritic cell; TCR, T cell receptor.
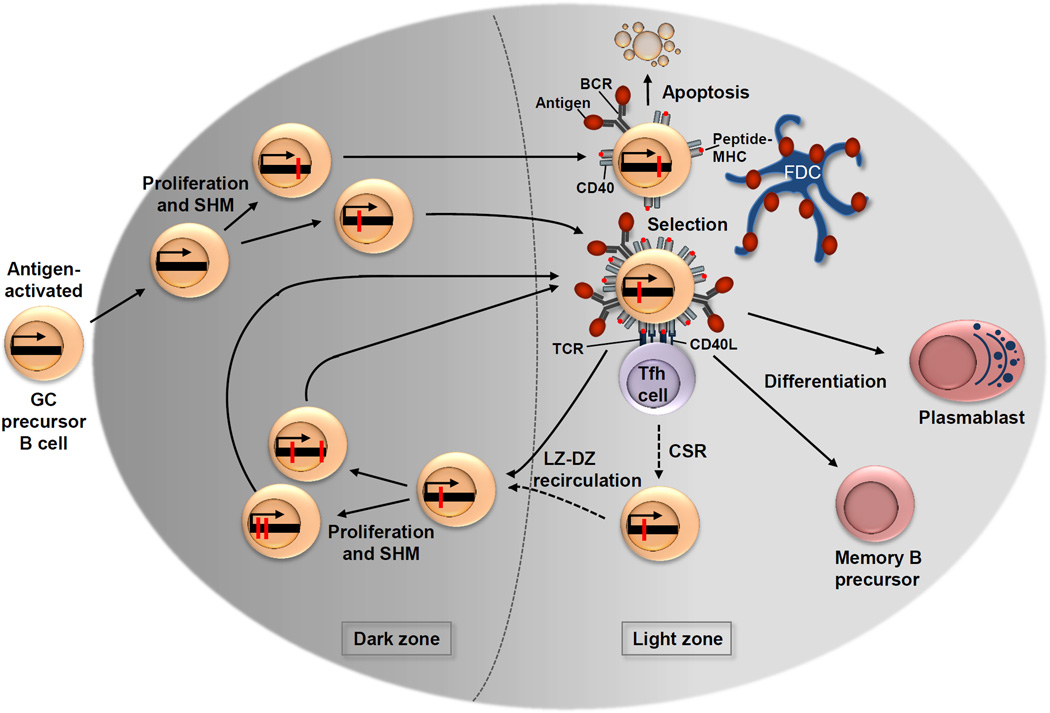
Antigen-activated germinal centre (GC) precursor B cells form the early GC at day 4, in which they differentiate into blasts that over the following days undergo clonal expansion until the mature GC that is characterized by the dark zone and the light zone forms at day 7. During proliferation, the process of somatic hypermutation (SHM) introduces base-pair changes into the V(D)J region of the rearranged immunoglobulin variable region (IgV) genes (red dots); some of these base-pair mutations lead to a change in the amino acid sequence. Dark zone B cells then move to the light zone, where the modified B cell receptor (BCR) of the light zone B cell, with help from immune cells, including T follicular helper cells (TFH cells) and follicular dendritic cells (FDCs), is selected for improved binding to the immunizing antigen. Among the newly generated light zone B cells that express BCR mutants resulting from SHM in the dark zone, higher BCR affinity is directly associated with greater antigen capture and leads to a higher density of peptide–MHC complexes presented on the surface of the B cell. This results in the greatest share of T cell help, which in turn drives positive selection. Therefore, newly generated light zone B cells that produce an unfavourable antibody are rendered unable to capture sufficient antigen and undergo apoptosis. Following positive selection, a subset of light zone B cells is instructed to recirculate to the dark zone. Light zone B cells may undergo immunoglobulin class-switch recombination (CSR) before light zone–dark zone recirculation, whereas other cells switch and directly differentiate (not depicted). Back in the dark zone, these cells undergo further proliferation and SHM, thus potentially generating antibody mutants with an improved affinity. Recirculation between the dark zone and the light zone facilitates several iterative rounds of mutation and selection, and within a short time, leads to the generation of high-affinity memory B cells and plasma cells. Antigen-selected light zone B cells eventually differentiate into memory B cell precursor cells and plasmablasts, which are the precursors of plasma cells. CD40L, CD40 ligand; TCR, T cell receptor.
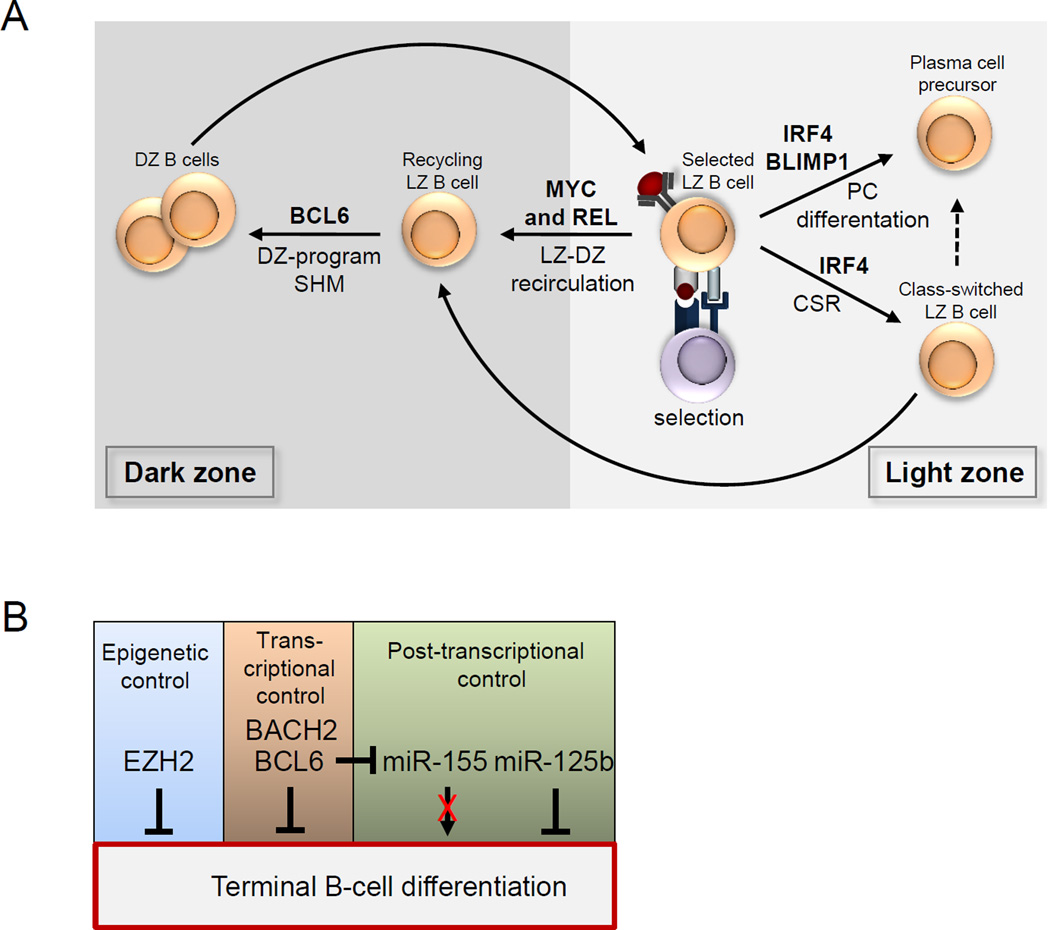
a Roles of transcriptional regulators in stages of germinal centre (GC) B cell development, as identified by genetic analysis, are shown. B cell lymphoma 6 (BCL-6) is required for the establishment of the specialized dark zone-associated gene expression programme that allows for somatic hypermutation (SHM) of the B cell receptor (BCR). MYC and REL are required for recirculation between the light zone and the dark zone, and thus for the maintenance of the GC reaction. Interferon-regulatory factor 4 (IRF4), although expressed in light zone B cells and associated with the downregulation of the dark zone-associated programme, is not required for GC maintenance. Instead, IRF4 is required for immunoglobulin class-switch recombination (CSR), and thus the frequency of isotype-switched B cells exiting the GC, and also for the production of GC-derived plasma cells. B lymphocyte-induced maturation protein 1 (BLIMP1) is also required for plasma cell differentiation. b Different genetic mechanisms maintain the dark zone phenotype by inhibiting terminal differentiation. The core catalytic component of polycomb repressive complex 2, enhancer of zeste homologue 2 (EZH2), inhibits terminal differentiation by silencing the expression of the plasma cell regulators Irf4 and Prdm1 (which encodes BLIMP1). BACH2 (BTB and CNC homologue 2) cooperates with BCL-6 and represses genes involved in post-GC differentiation. BCL-6 transcriptionally represses plasma cell-associated genes including Prdm1, directly and indirectly via downregulation of the microRNA miR-155 that is associated with terminating the dark zone-associated programme. miR-125b downregulates the expression of IRF4 and BLIMP1; as this microRNA is highly expressed in dark zone B cells, it probably contributes to the repression of expression of Irf4 and Prdm1. CD40L, CD40 ligand; TCR, T cell receptor.
References
-
- MacLennan IC. Germinal centers. Annu. Rev. Immunol. 1994;12:117–139. - PubMed
-
- Jacob J, Kelsoe G, Rajewsky K, Weiss U. Intraclonal generation of antibody mutants in germinal centres. Nature. 1991;354:389–392. - PubMed
-
- Berek C, Berger A, Apel M. Maturation of the immune response in germinal centers. Cell. 1991;67:1121–1129. - PubMed
-
- Victora GD, Nussenzweig MC. Germinal centers. Annu. Rev. Immunol. 2012;30:429–457. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
