

The motility of axonemal dynein is regulated by the tubulin code
- PMID: 25658008
- PMCID: PMC4808650
- DOI: 10.1016/j.bpj.2014.10.061
The motility of axonemal dynein is regulated by the tubulin code
Abstract
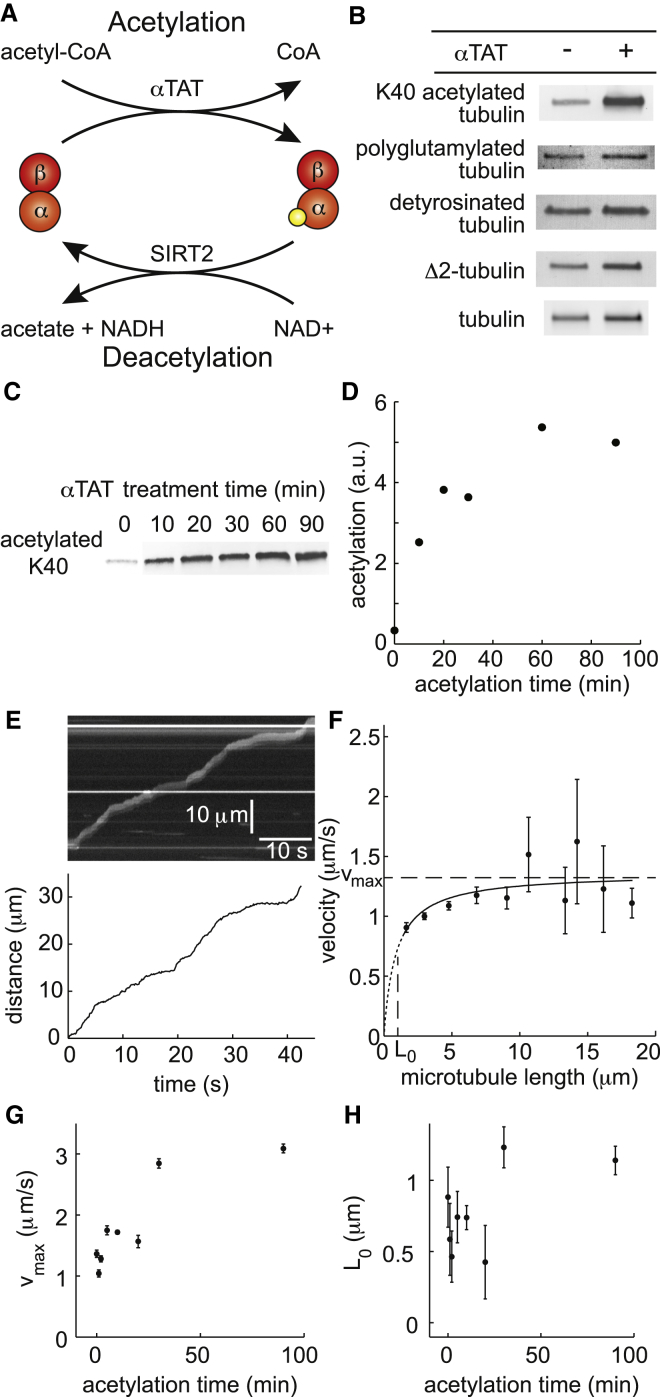
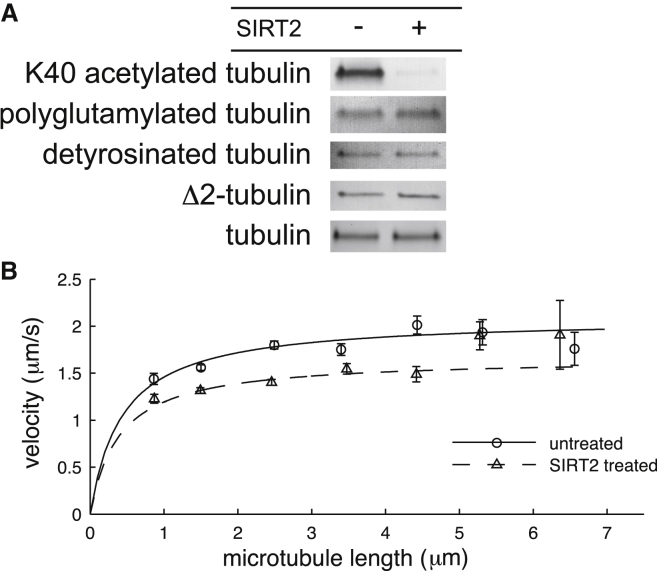
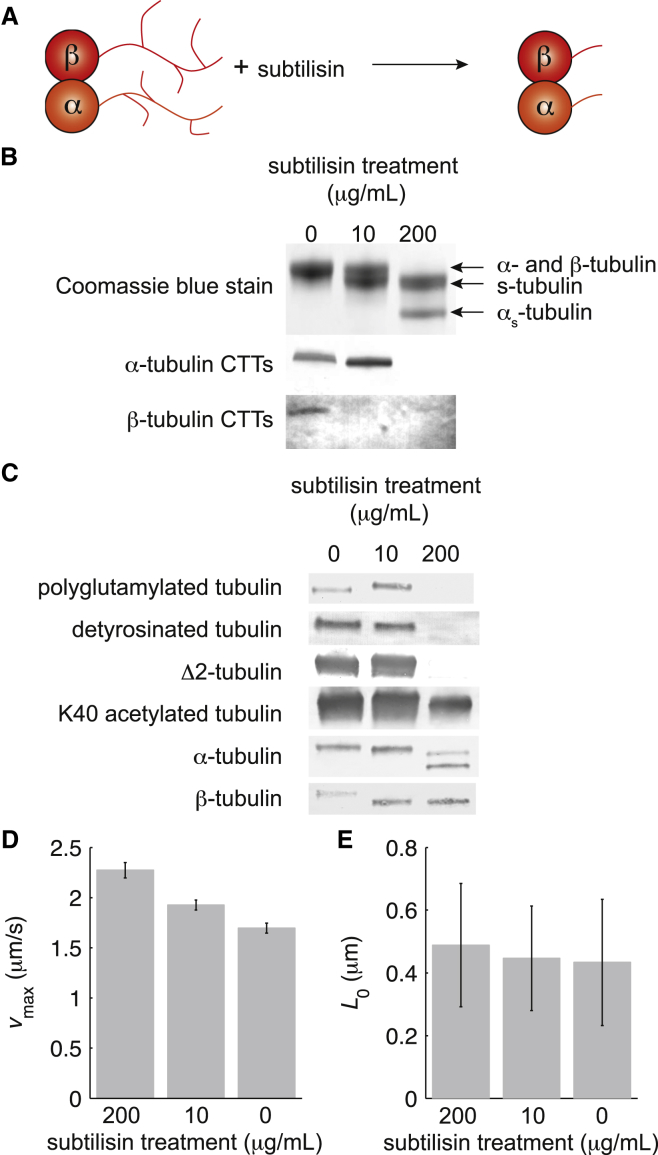
Microtubule diversity, arising from the utilization of different tubulin genes and from posttranslational modifications, regulates many cellular processes including cell division, neuronal differentiation and growth, and centriole assembly. In the case of cilia and flagella, multiple cell biological studies show that microtubule diversity is important for axonemal assembly and motility. However, it is not known whether microtubule diversity directly influences the activity of the axonemal dyneins, the motors that drive the beating of the axoneme, nor whether the effects on motility are indirect, perhaps through regulatory pathways upstream of the motors, such as the central pair, radial spokes, or dynein regulatory complex. To test whether microtubule diversity can directly regulate the activity of axonemal dyneins, we asked whether in vitro acetylation or deacetylation of lysine 40 (K40), a major posttranslational modification of α-tubulin, or whether proteolytic cleavage of the C-terminal tail (CTT) of α- and β-tubulin, the location of detyrosination, polyglutamylation, and polyglycylation modifications as well as most of the genetic diversity, can influence the activity of outer arm axonemal dynein in motility assays using purified proteins. By quantifying the motility with displacement-weighted velocity analysis and mathematically modeling the results, we found that K40 acetylation increases and CTTs decrease axonemal dynein motility. These results show that axonemal dynein directly deciphers the tubulin code, which has important implications for eukaryotic ciliary beat regulation.
Figures
References
-
- Verhey K.J., Gaertig J. The tubulin code. Cell Cycle. 2007;6:2152–2160. - PubMed
-
- Ludueña R.F. Multiple forms of tubulin: different gene products and covalent modifications. Int. Rev. Cytol. 1998;178:207–275. - PubMed
-
- Janke C., Bulinski J.C. Post-translational regulation of the microtubule cytoskeleton: mechanisms and functions. Nat. Rev. Mol. Cell Biol. 2011;12:773–786. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
