

The host-protective effect of arabinosylated lipoarabinomannan against Leishmania donovani infection is associated with restoration of IFN-γ responsiveness
- PMID: 25658110
- PMCID: PMC4319725
- DOI: 10.1371/journal.pone.0117247
The host-protective effect of arabinosylated lipoarabinomannan against Leishmania donovani infection is associated with restoration of IFN-γ responsiveness
Abstract
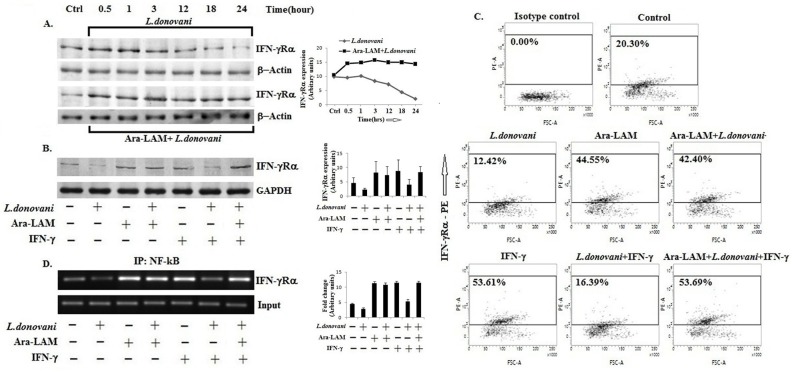
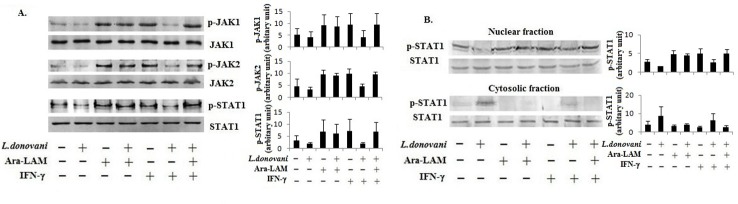
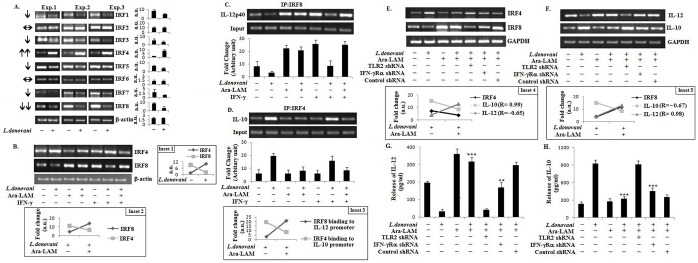
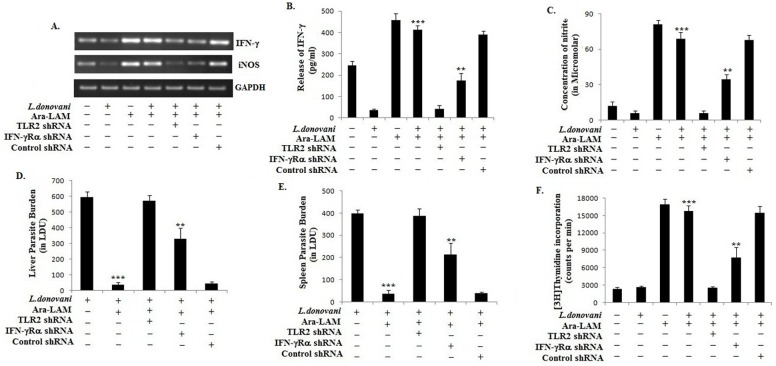
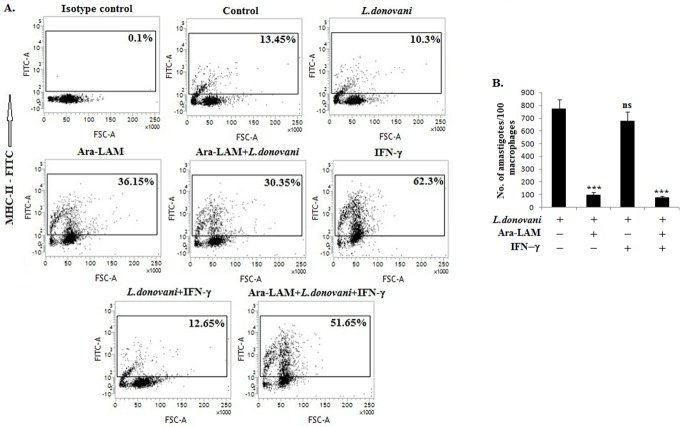
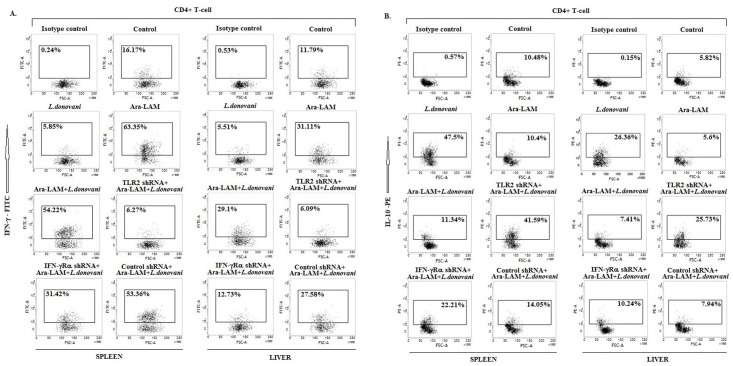
Visceral leishmaniasis (VL), which is endemic as a major infectious disease in the tropical and subtropical countries, is caused by a protozoan parasite Leishmania donovani. At present, restricted treatment options and lack of vaccines intensify the problem of controlling VL. Therefore, finding a novel immunoprophylactic or therapeutic principle is a pressing need. Here, we report that arabinosylated lipoarabinomannan (Ara-LAM), a TLR2-ligand isolated from Mycobacterium smegmatis, exhibits a strong immunomodulatory property that conferred protection against L. donovani infection. Although, Ara-LAM modulates TLR2 and MAPK signaling, it is not known whether Ara-LAM involves IFN-γ signaling for effective parasite clearance. Because, it is reported that IFN-γ signaling, a principle mediator of NO generation and macrophage and Tcell activation, is hampered during leishmanial pathogenesis. Ara-LAM increases IFN-γ receptor expression and potentiates IFN-γ receptor signaling through JAK-STAT pathway. Moreover, Ara-LAM reciprocally modulates IRF4 and IRF8 expression and reinstates anti-leishmanial Th1 response that eventuates in significantly reduced parasite load in spleen and liver of L. donovani-infected BALB/c mice. IFN-γRα silencing resulted in the suppression of these host-protective mechanisms affected by Ara-LAM. Thus, Ara-LAM-mediated restoration of IFN-γ responsiveness is a novel immuno-modulatory principle for protection against L. donovani susceptible host.
Conflict of interest statement
Figures
Similar articles
-
Immunomodulation of host-protective immune response by regulating Foxp3 expression and Treg function in Leishmania-infected BALB/c mice: critical role of IRF1.Pathog Dis. 2015 Nov;73(8):ftv063. doi: 10.1093/femspd/ftv063. Epub 2015 Aug 21. Pathog Dis. 2015. PMID: 26297915 Free PMC article.
-
Arabinosylated lipoarabinomannan skews Th2 phenotype towards Th1 during Leishmania infection by chromatin modification: involvement of MAPK signaling.PLoS One. 2011;6(9):e24141. doi: 10.1371/journal.pone.0024141. Epub 2011 Sep 14. PLoS One. 2011. PMID: 21935379 Free PMC article.
-
Immunomodulatory role of arabinosylated lipoarabinomannan on Leishmania donovani infected murine macrophages.Indian J Biochem Biophys. 2007 Oct;44(5):366-72. Indian J Biochem Biophys. 2007. PMID: 18341212
-
Leishmania intercepts IFN-γR signaling at multiple levels in macrophages.Cytokine. 2022 Sep;157:155956. doi: 10.1016/j.cyto.2022.155956. Epub 2022 Jul 1. Cytokine. 2022. PMID: 35785668 Review.
-
Exploitation of host cell signaling machinery: activation of macrophage phosphotyrosine phosphatases as a novel mechanism of molecular microbial pathogenesis.J Leukoc Biol. 2000 Apr;67(4):464-70. doi: 10.1002/jlb.67.4.464. J Leukoc Biol. 2000. PMID: 10770277 Review.
Cited by
-
Toll-Like Receptor 2 Is Required for Inflammatory Process Development during Leishmania infantum Infection.Front Microbiol. 2017 Feb 23;8:262. doi: 10.3389/fmicb.2017.00262. eCollection 2017. Front Microbiol. 2017. PMID: 28280488 Free PMC article.
-
Immunomodulation of host-protective immune response by regulating Foxp3 expression and Treg function in Leishmania-infected BALB/c mice: critical role of IRF1.Pathog Dis. 2015 Nov;73(8):ftv063. doi: 10.1093/femspd/ftv063. Epub 2015 Aug 21. Pathog Dis. 2015. PMID: 26297915 Free PMC article.
-
Paradoxical immune response in leishmaniasis: The role of toll-like receptors in disease progression.Parasite Immunol. 2022 Apr;44(4-5):e12910. doi: 10.1111/pim.12910. Epub 2022 Feb 28. Parasite Immunol. 2022. PMID: 35119120 Free PMC article. Review.
-
The Mitogen-Activated Protein Kinase (MAPK) Pathway: Role in Immune Evasion by Trypanosomatids.Front Microbiol. 2016 Feb 24;7:183. doi: 10.3389/fmicb.2016.00183. eCollection 2016. Front Microbiol. 2016. PMID: 26941717 Free PMC article. Review.
References
-
- Chappuis F, Sundar S, Hailu A, Ghalib H, Rijal S, et al. (2007) Visceral leishmaniasis: what are the needs for diagnosis, treatment and control. Nat Rev Microbiol 5: 873–882. - PubMed
-
- Vanaerschot M, Dumetz F, Roy S, Ponte-Sucre A, Arevalo J, et al. (2014) Treatment failure in leishmaniasis: drug-resistance or another (epi-) phenotype? Expert Rev Anti Infect Ther [Epub ahead of print] PubMed PMID: 24802998. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
