

Essential domains of Anaplasma phagocytophilum invasins utilized to infect mammalian host cells
- PMID: 25658707
- PMCID: PMC4450072
- DOI: 10.1371/journal.ppat.1004669
Essential domains of Anaplasma phagocytophilum invasins utilized to infect mammalian host cells
Abstract
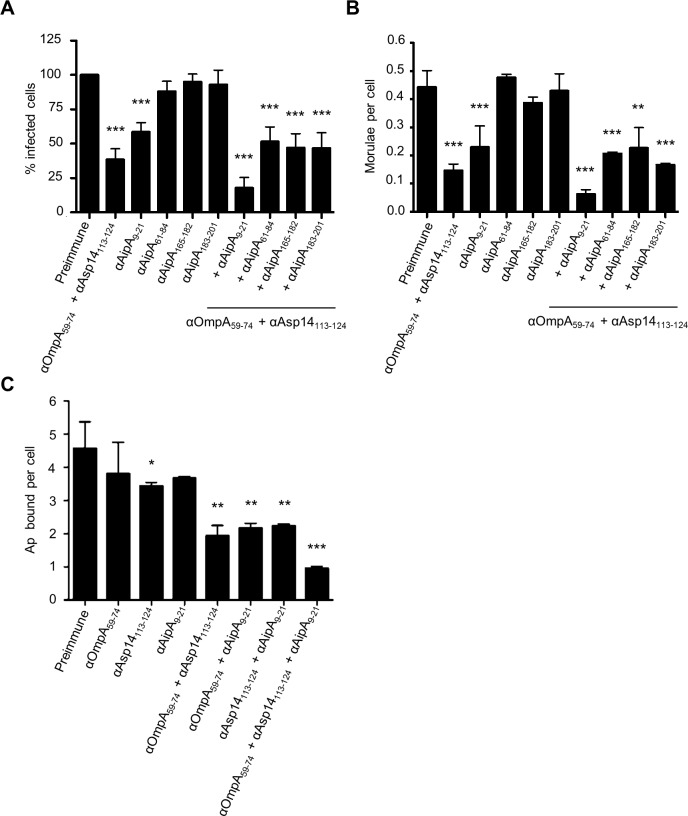
Anaplasma phagocytophilum causes granulocytic anaplasmosis, an emerging disease of humans and domestic animals. The obligate intracellular bacterium uses its invasins OmpA, Asp14, and AipA to infect myeloid and non-phagocytic cells. Identifying the domains of these proteins that mediate binding and entry, and determining the molecular basis of their interactions with host cell receptors would significantly advance understanding of A. phagocytophilum infection. Here, we identified the OmpA binding domain as residues 59 to 74. Polyclonal antibody generated against a peptide spanning OmpA residues 59 to 74 inhibited A. phagocytophilum infection of host cells and binding to its receptor, sialyl Lewis x (sLe(x)-capped P-selectin glycoprotein ligand 1. Molecular docking analyses predicted that OmpA residues G61 and K64 interact with the two sLe(x) sugars that are important for infection, α2,3-sialic acid and α1,3-fucose. Amino acid substitution analyses demonstrated that K64 was necessary, and G61 was contributory, for recombinant OmpA to bind to host cells and competitively inhibit A. phagocytophilum infection. Adherence of OmpA to RF/6A endothelial cells, which express little to no sLe(x) but express the structurally similar glycan, 6-sulfo-sLe(x), required α2,3-sialic acid and α1,3-fucose and was antagonized by 6-sulfo-sLe(x) antibody. Binding and uptake of OmpA-coated latex beads by myeloid cells was sensitive to sialidase, fucosidase, and sLe(x) antibody. The Asp14 binding domain was also defined, as antibody specific for residues 113 to 124 inhibited infection. Because OmpA, Asp14, and AipA each contribute to the infection process, it was rationalized that the most effective blocking approach would target all three. An antibody cocktail targeting the OmpA, Asp14, and AipA binding domains neutralized A. phagocytophilum binding and infection of host cells. This study dissects OmpA-receptor interactions and demonstrates the effectiveness of binding domain-specific antibodies for blocking A. phagocytophilum infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures








References
-
- Hopkins RS, Jajosky RA, Hall PA, Adams DA, Connor FJ, et al. (2005) Summary of notifiable diseases—United States, 2003. MMWR Morb Mortal Wkly Rep 52: 1–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
