

FANCD2, FANCJ and BRCA2 cooperate to promote replication fork recovery independently of the Fanconi Anemia core complex
- PMID: 25659033
- PMCID: PMC4614542
- DOI: 10.4161/15384101.2014.987614
FANCD2, FANCJ and BRCA2 cooperate to promote replication fork recovery independently of the Fanconi Anemia core complex
Abstract
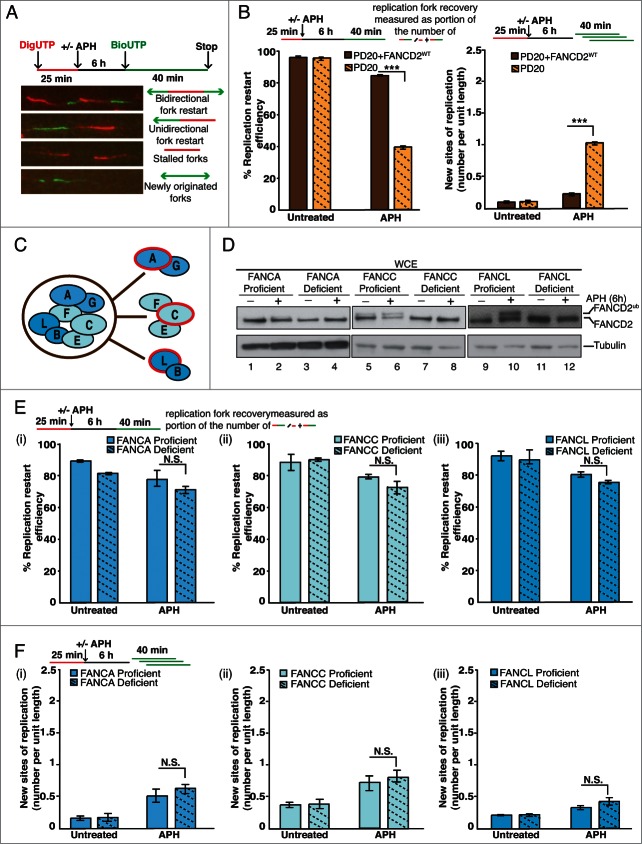
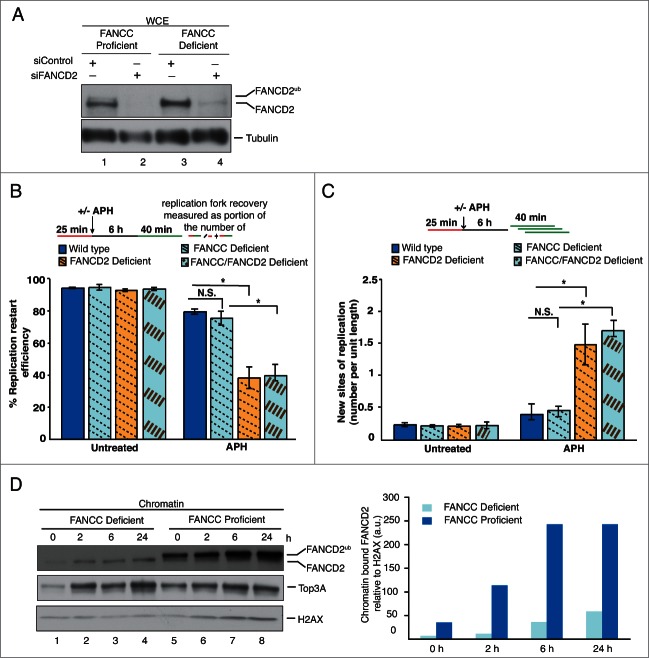
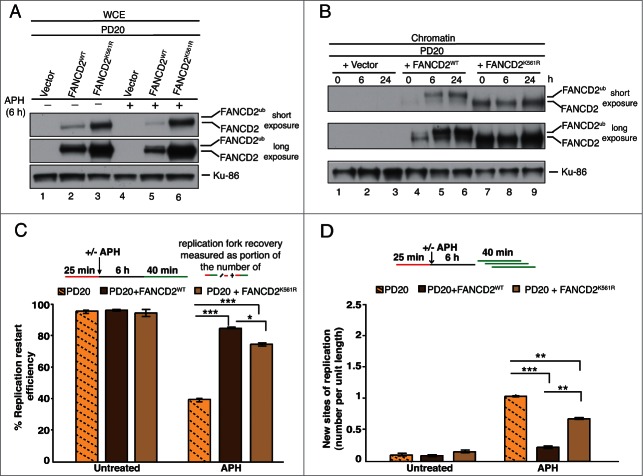
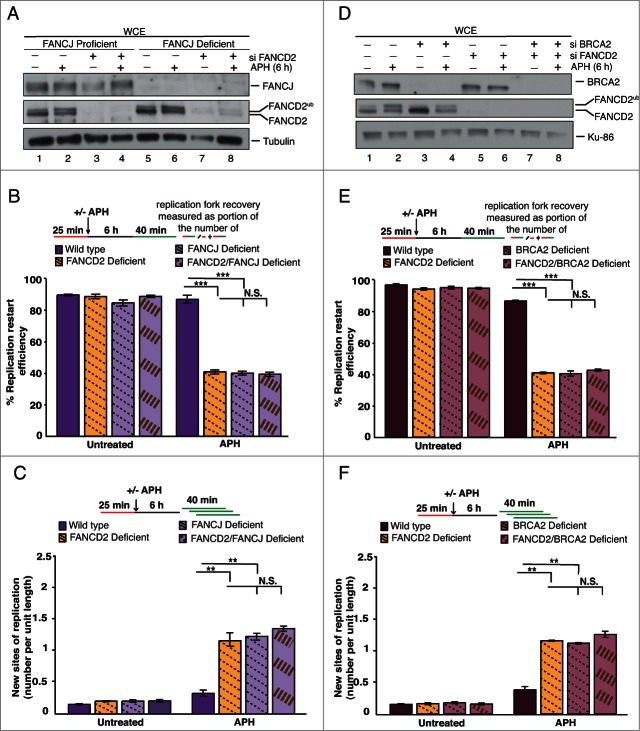
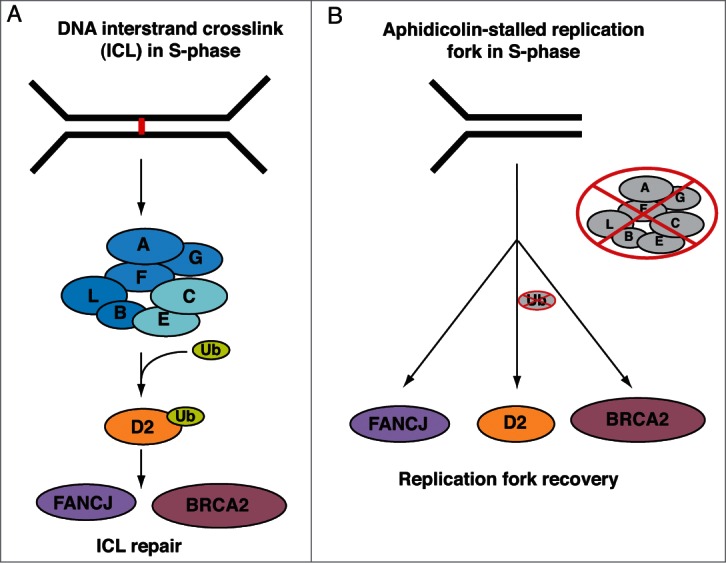
Fanconi Anemia (FA) is an inherited multi-gene cancer predisposition syndrome that is characterized on the cellular level by a hypersensitivity to DNA interstrand crosslinks (ICLs). To repair these lesions, the FA pathway proteins are thought to act in a linear hierarchy: Following ICL detection, an upstream FA core complex monoubiquitinates the central FA pathway members FANCD2 and FANCI, followed by their recruitment to chromatin. Chromatin-bound monoubiquitinated FANCD2 and FANCI subsequently coordinate DNA repair factors including the downstream FA pathway members FANCJ and FANCD1/BRCA2 to repair the DNA ICL. Importantly, we recently showed that FANCD2 has additional independent roles: it binds chromatin and acts in concert with the BLM helicase complex to promote the restart of aphidicolin (APH)-stalled replication forks, while suppressing the firing of new replication origins. Here, we show that FANCD2 fulfills these roles independently of the FA core complex-mediated monoubiquitination step. Following APH treatment, nonubiquitinated FANCD2 accumulates on chromatin, recruits the BLM complex, and promotes robust replication fork recovery regardless of the absence or presence of a functional FA core complex. In contrast, the downstream FA pathway members FANCJ and BRCA2 share FANCD2's role in replication fork restart and the suppression of new origin firing. Our results support a non-linear FA pathway model at stalled replication forks, where the nonubiquitinated FANCD2 isoform - in concert with FANCJ and BRCA2 - fulfills a specific function in promoting efficient replication fork recovery independently of the FA core complex.
Keywords: FA pathway; FANCD1/BRCA2; FANCD2; FANCD2 monoubiquitination; FANCJ; Fanconi Anemia; replication fork recovery.
Figures
References
-
- Kee Y, D'Andrea AD. Molecular pathogenesis and clinical management of Fanconi anemia. J Clin Invest 2012; 122:3799-806; PMID:23114602; http://dx.doi.org/ 10.1172/JCI58321 - DOI - PMC - PubMed
-
- Naim V, Rosselli F. The FANC pathway and BLM collaborate during mitosis to prevent micro-nucleation and chromosome abnormalities. Nat Cell Biol 2009; 11:761-8; PMID:19465921; http://dx.doi.org/ 10.1038/ncb1883 - DOI - PubMed
-
- Kee Y, D'Andrea AD. Expanded roles of the Fanconi anemia pathway in preserving genomic stability. Genes Dev 2010; 24:1680-94; PMID:20713514; http://dx.doi.org/ 10.1101/gad.1955310 - DOI - PMC - PubMed
-
- Schlacher K, Wu H, Jasin M. A distinct replication fork protection pathway connects fanconi anemia tumor suppressors to RAD51-BRCA1/2. Cancer Cell 2012; 22:106-16; PMID:22789542; http://dx.doi.org/ 10.1016/j.ccr.2012.05.015 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous