

Identification of G protein-biased agonists that fail to recruit β-arrestin or promote internalization of the D1 dopamine receptor
- PMID: 25660762
- PMCID: PMC5234767
- DOI: 10.1021/acschemneuro.5b00020
Identification of G protein-biased agonists that fail to recruit β-arrestin or promote internalization of the D1 dopamine receptor
Abstract
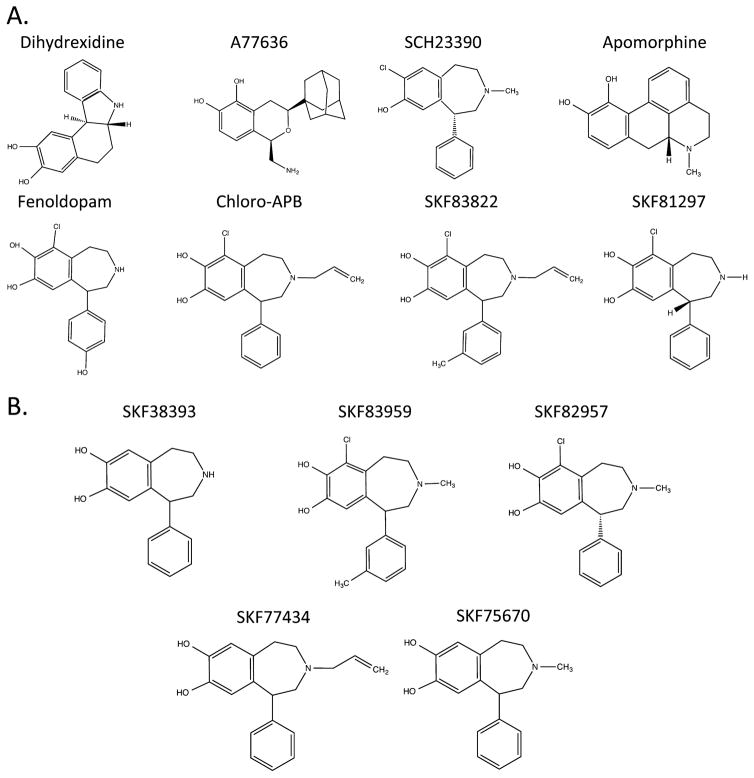
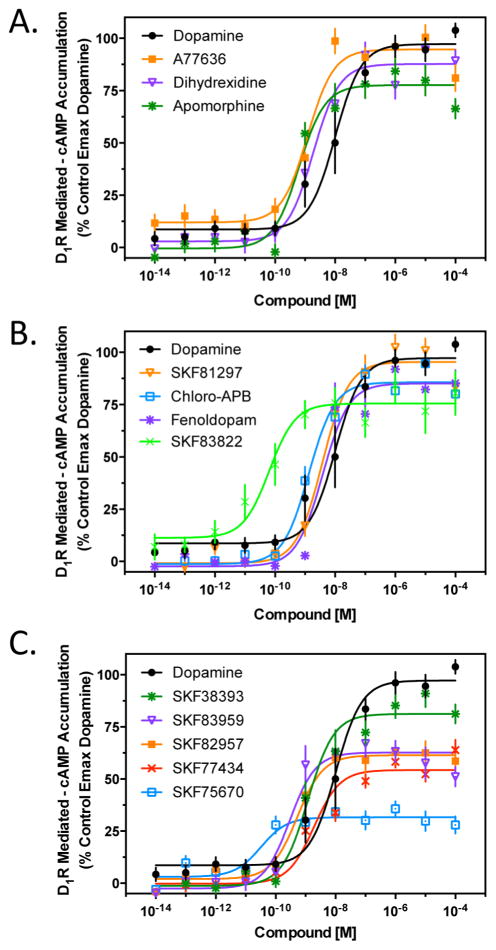
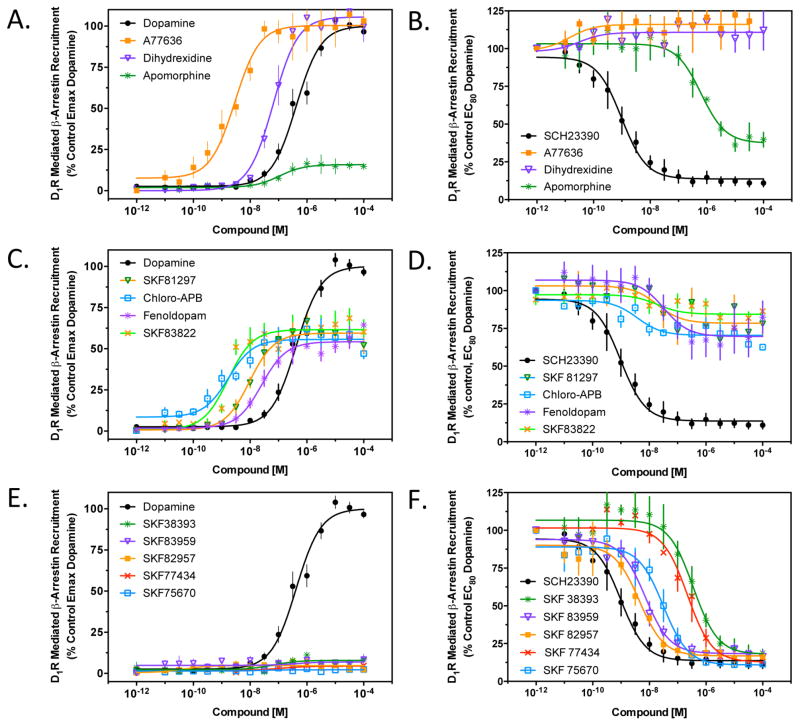
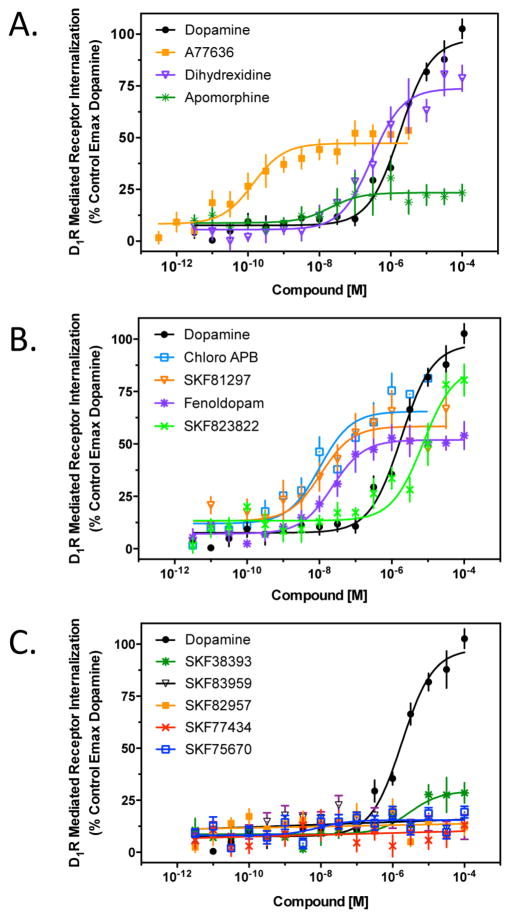
The D1 dopamine receptor (D1R) has been implicated in numerous neuropsychiatric disorders, and D1R-selective ligands have potential as therapeutic agents. Previous studies have identified substituted benzazepines as D1R-selective agonists, but the in vivo effects of these compounds have not correlated well with their in vitro pharmacological activities. A series of substituted benzazepines, and structurally dissimilar D1R-selective agonists, were tested for their functional effects on D1R-mediated cAMP accumulation, D1R-promoted β-arrestin recruitment, and D1R internalization using live cell functional assays. All compounds tested elicited an increase in the level of cAMP accumulation, albeit with a range of efficacies. However, when the compounds were evaluated for β-arrestin recruitment, a subset of substituted benzazepines, SKF83959, SKF38393, SKF82957, SKF77434, and SKF75670, failed to activate this pathway, whereas the others showed similar activation efficacies as seen with cAMP accumulation. When tested as antagonists, the five biased compounds all inhibited dopamine-stimulated β-arrestin recruitment. Further, D1R internalization assays revealed a corroborating pattern of activity in that the G protein-biased compounds failed to promote D1R internalization. Interestingly, the biased signaling was unique for the D1R, as the same compounds were agonists of the related D5 dopamine receptor (D5R), but revealed no signaling bias. We have identified a group of substituted benzazepine ligands that are agonists at D1R-mediated G protein signaling, but antagonists of D1R recruitment of β-arrestin, and also devoid of agonist-induced receptor endocytosis. These data may be useful for interpreting the contrasting effects of these compounds in vitro versus in vivo, and also for the understanding of pathway-selective signaling of the D1R.
Keywords: D1 receptor; Dopamine; G protein; benzazepine; biased agonism; functional selectivity; β-arrestin.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
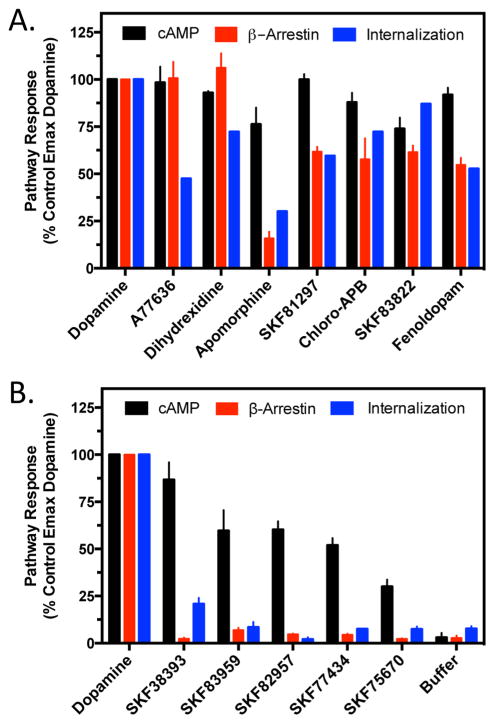
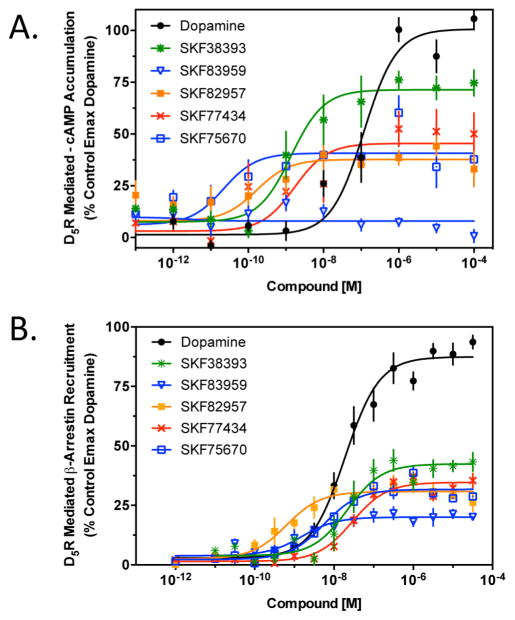
References
-
- Rankin ML, Hazelwood LA, Free RB, Namkung Y, Rex EB, Roof RA, Sibley DR. Molecular pharmacology of the dopamine receptors. In: Iversen LL, Iversen SD, Dunnett SB, Bjorklund A, editors. Dopamine Handbook. Oxford University Press; New York: 2009. pp. 63–87.
-
- Kapur S, Mamo D. Half a century of antipsychotics and still a central role for dopamine D2 receptors. Prog Neuro-Psychopharmacol Biol Psychiatry. 2003;27:1081–1090. - PubMed
-
- Sibley DR, Monsma FJ., Jr Molecular biology of dopamine receptors. Trends Pharmacol Sci. 1992;13:61–69. - PubMed
-
- Beaulieu JM, Gainetdinov RR. The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol Rev. 2011;63:182–217. - PubMed
-
- Swinney DC. Successful drug discovery. Curr Top Med Chem. 2006;6:403–404. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
