

Molecular mechanism and evolution of guanylate kinase regulation by (p)ppGpp
- PMID: 25661490
- PMCID: PMC4336630
- DOI: 10.1016/j.molcel.2014.12.037
Molecular mechanism and evolution of guanylate kinase regulation by (p)ppGpp
Abstract
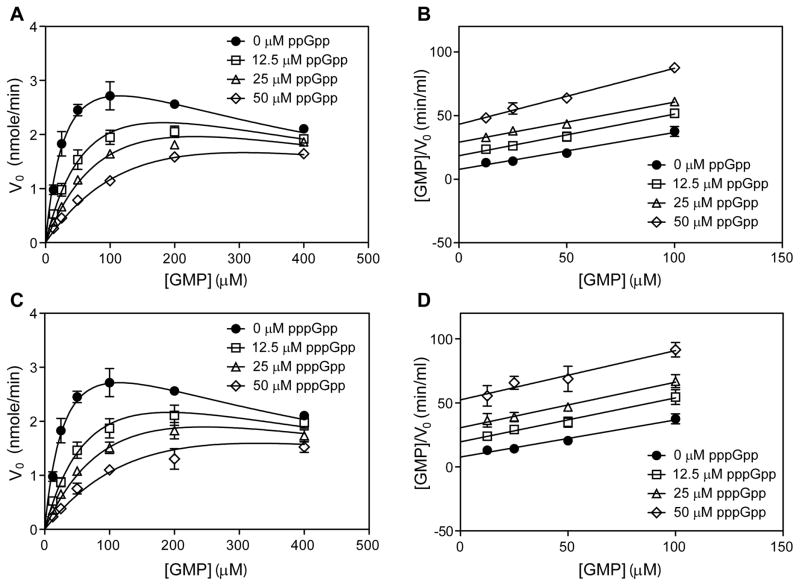
The nucleotide (p)ppGpp mediates bacterial stress responses, but its targets and underlying mechanisms of action vary among bacterial species and remain incompletely understood. Here, we characterize the molecular interaction between (p)ppGpp and guanylate kinase (GMK), revealing the importance of this interaction in adaptation to starvation. Combining structural and kinetic analyses, we show that (p)ppGpp binds the GMK active site and competitively inhibits the enzyme. The (p)ppGpp-GMK interaction prevents the conversion of GMP to GDP, resulting in GMP accumulation upon amino acid downshift. Abolishing this interaction leads to excess (p)ppGpp and defective adaptation to amino acid starvation. A survey of GMKs from phylogenetically diverse bacteria shows that the (p)ppGpp-GMK interaction is conserved in members of Firmicutes, Actinobacteria, and Deinococcus-Thermus, but not in Proteobacteria, where (p)ppGpp regulates RNA polymerase (RNAP). We propose that GMK is an ancestral (p)ppGpp target and RNAP evolved more recently as a direct target in Proteobacteria.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures






Similar articles
-
From (p)ppGpp to (pp)pGpp: Characterization of Regulatory Effects of pGpp Synthesized by the Small Alarmone Synthetase of Enterococcus faecalis.J Bacteriol. 2015 Sep;197(18):2908-19. doi: 10.1128/JB.00324-15. Epub 2015 Jun 29. J Bacteriol. 2015. PMID: 26124242 Free PMC article.
-
Differential binding of ppGpp and pppGpp to E. coli RNA polymerase: photo-labeling and mass spectral studies.Genes Cells. 2015 Dec;20(12):1006-16. doi: 10.1111/gtc.12304. Epub 2015 Oct 8. Genes Cells. 2015. PMID: 26606426
-
Evolution of (p)ppGpp-HPRT regulation through diversification of an allosteric oligomeric interaction.Elife. 2019 Sep 25;8:e47534. doi: 10.7554/eLife.47534. Elife. 2019. PMID: 31552824 Free PMC article.
-
The magic dance of the alarmones (p)ppGpp.Mol Microbiol. 2016 Aug;101(4):531-44. doi: 10.1111/mmi.13412. Epub 2016 Jun 10. Mol Microbiol. 2016. PMID: 27149325 Review.
-
Direct binding targets of the stringent response alarmone (p)ppGpp.Mol Microbiol. 2012 Sep;85(6):1029-43. doi: 10.1111/j.1365-2958.2012.08177.x. Epub 2012 Aug 2. Mol Microbiol. 2012. PMID: 22812515 Review.
Cited by
-
Diverse molecular mechanisms of transcription regulation by the bacterial alarmone ppGpp.Mol Microbiol. 2022 Feb;117(2):252-260. doi: 10.1111/mmi.14860. Epub 2021 Dec 25. Mol Microbiol. 2022. PMID: 34894005 Free PMC article. Review.
-
Novel (p)ppGpp Binding and Metabolizing Proteins of Escherichia coli.mBio. 2018 Mar 6;9(2):e02188-17. doi: 10.1128/mBio.02188-17. mBio. 2018. PMID: 29511080 Free PMC article.
-
ppGpp negatively impacts ribosome assembly affecting growth and antimicrobial tolerance in Gram-positive bacteria.Proc Natl Acad Sci U S A. 2016 Mar 22;113(12):E1710-9. doi: 10.1073/pnas.1522179113. Epub 2016 Mar 7. Proc Natl Acad Sci U S A. 2016. PMID: 26951678 Free PMC article.
-
From (p)ppGpp to (pp)pGpp: Characterization of Regulatory Effects of pGpp Synthesized by the Small Alarmone Synthetase of Enterococcus faecalis.J Bacteriol. 2015 Sep;197(18):2908-19. doi: 10.1128/JB.00324-15. Epub 2015 Jun 29. J Bacteriol. 2015. PMID: 26124242 Free PMC article.
-
Phenolic Amides Are Potent Inhibitors of De Novo Nucleotide Biosynthesis.Appl Environ Microbiol. 2015 Sep 1;81(17):5761-72. doi: 10.1128/AEM.01324-15. Epub 2015 Jun 12. Appl Environ Microbiol. 2015. PMID: 26070680 Free PMC article.
References
-
- Bennett HJ, Pearce DM, Glenn S, Taylor CM, Kuhn M, Sonenshein AL, Andrew PW, Roberts IS. Characterization of relA and codY mutants of Listeria monocytogenes: identification of the CodY regulon and its role in virulence. Mol Microbiol. 2007;63:1453–1467. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
