

A Big Bang model of human colorectal tumor growth
- PMID: 25665006
- PMCID: PMC4575589
- DOI: 10.1038/ng.3214
A Big Bang model of human colorectal tumor growth
Abstract
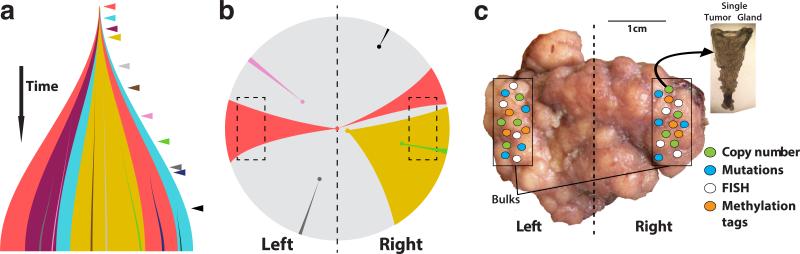
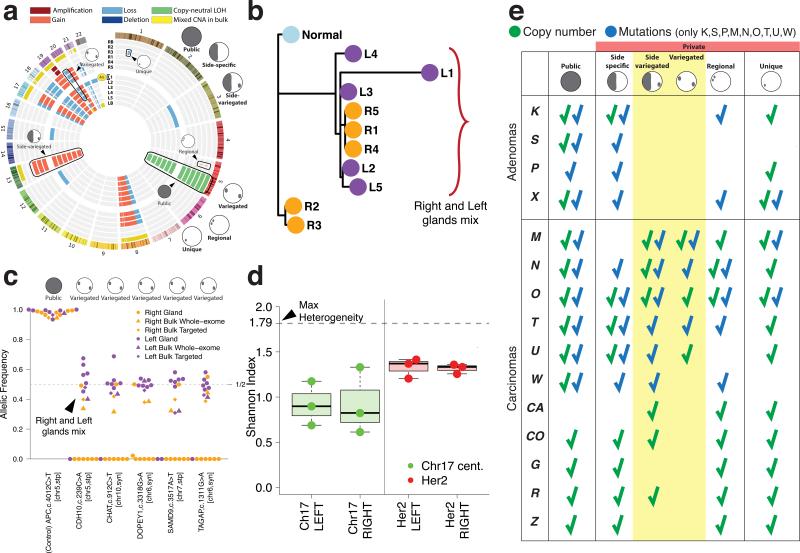
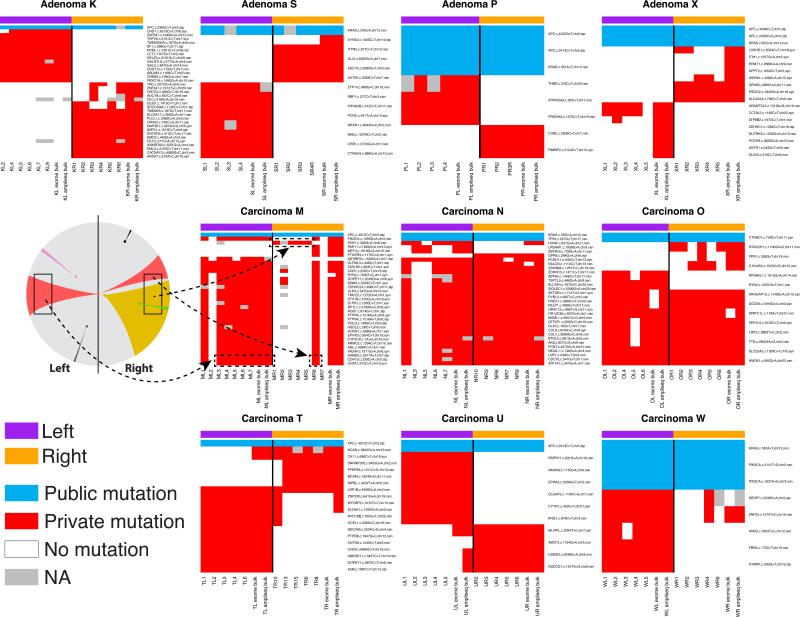
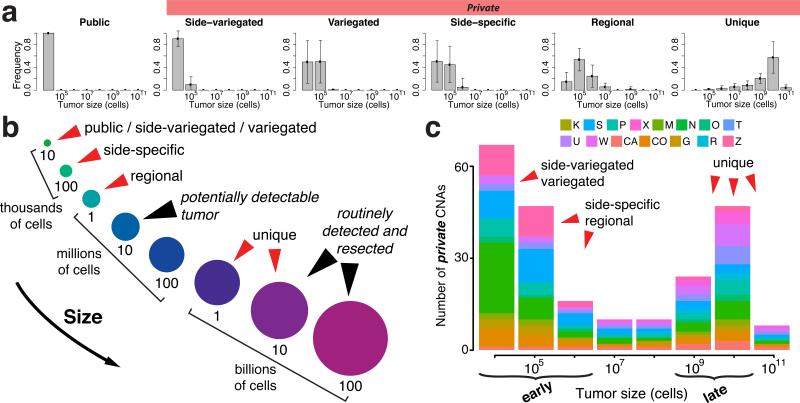
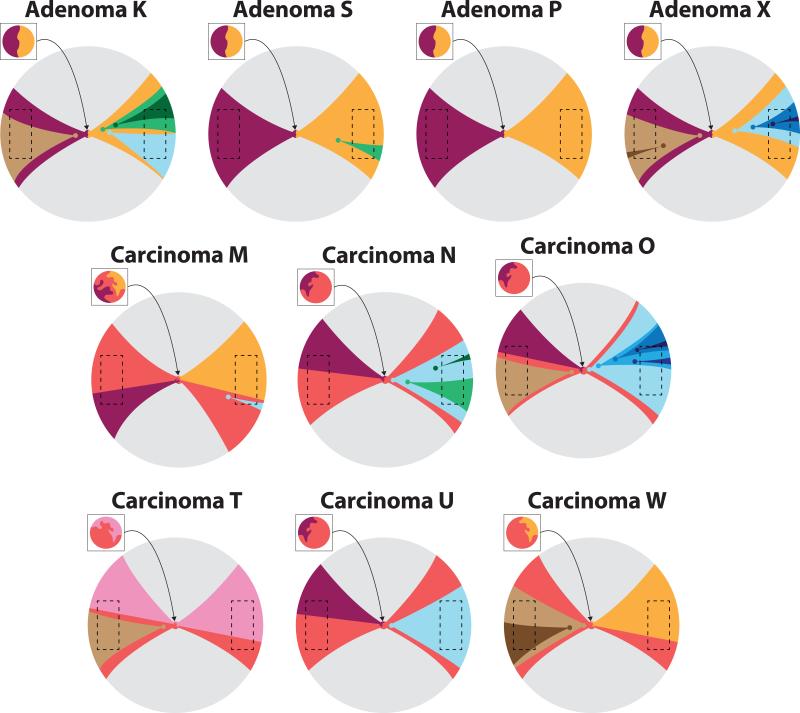
What happens in early, still undetectable human malignancies is unknown because direct observations are impractical. Here we present and validate a 'Big Bang' model, whereby tumors grow predominantly as a single expansion producing numerous intermixed subclones that are not subject to stringent selection and where both public (clonal) and most detectable private (subclonal) alterations arise early during growth. Genomic profiling of 349 individual glands from 15 colorectal tumors showed an absence of selective sweeps, uniformly high intratumoral heterogeneity (ITH) and subclone mixing in distant regions, as postulated by our model. We also verified the prediction that most detectable ITH originates from early private alterations and not from later clonal expansions, thus exposing the profile of the primordial tumor. Moreover, some tumors appear 'born to be bad', with subclone mixing indicative of early malignant potential. This new model provides a quantitative framework to interpret tumor growth dynamics and the origins of ITH, with important clinical implications.
Figures
Comment in
-
Big Bang and context-driven collapse.Nat Genet. 2015 Mar;47(3):196-7. doi: 10.1038/ng.3231. Nat Genet. 2015. PMID: 25711865
-
Heterogeneity: Explosive beginnings.Nat Rev Cancer. 2015 Apr;15(4):198-9. doi: 10.1038/nrc3928. Nat Rev Cancer. 2015. PMID: 25801613 No abstract available.
References
-
- Fearon ER, Fearon ER, Vogelstein B, Vogelstein B. A genetic model for colorectal tumorigenesis. Cell. 1990;61:759–767. - PubMed
Publication types
MeSH terms
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
