

Diagnosing oxidative stress in bacteria: not as easy as you might think
- PMID: 25666086
- PMCID: PMC4380616
- DOI: 10.1016/j.mib.2015.01.004
Diagnosing oxidative stress in bacteria: not as easy as you might think
Abstract
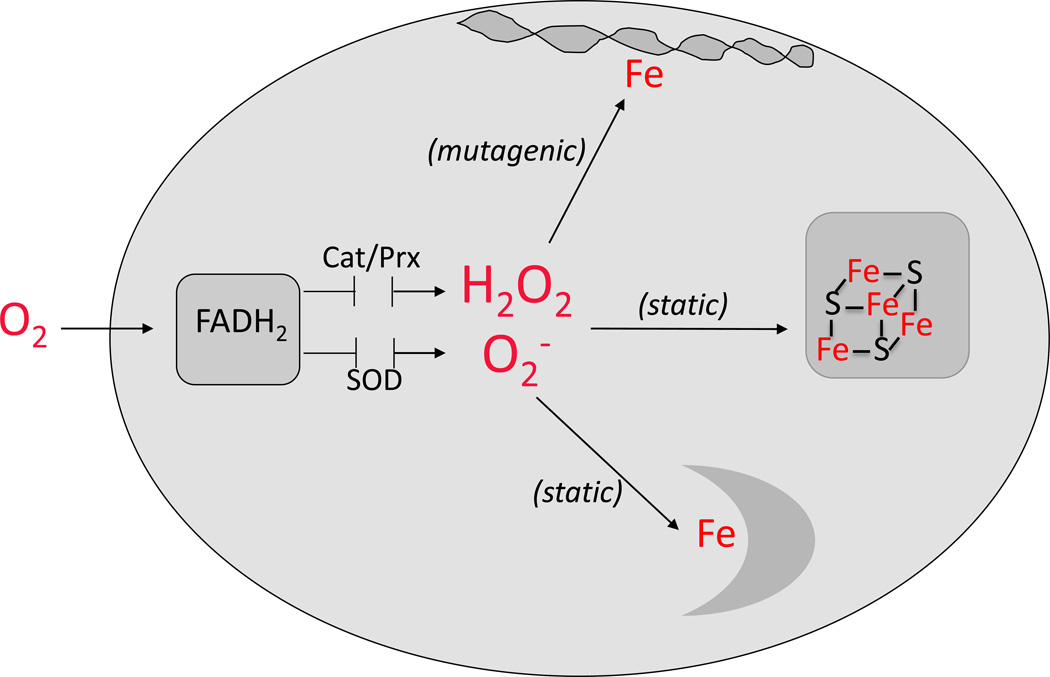
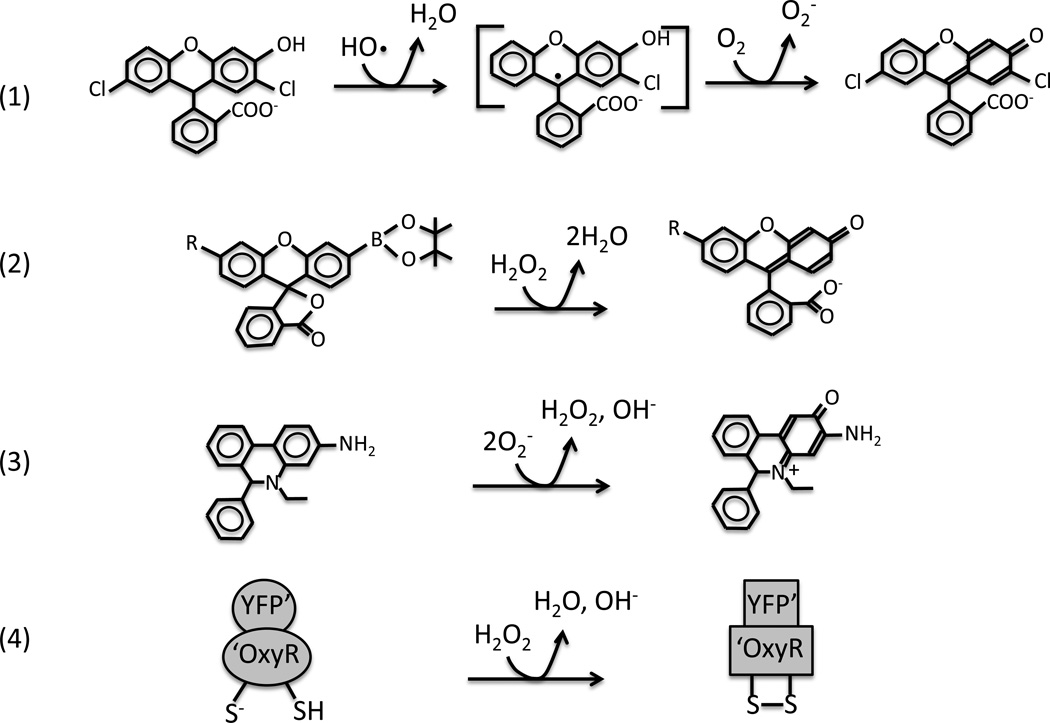
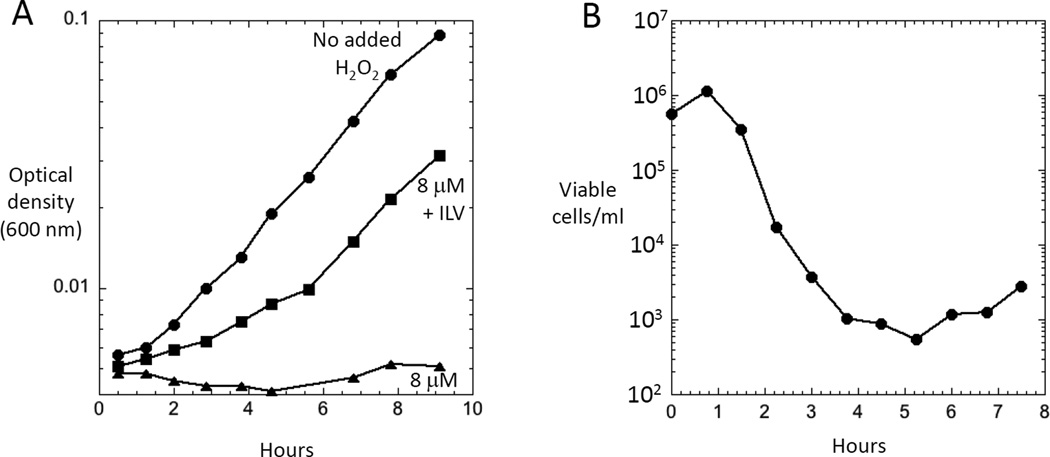
Microorganisms are vulnerable to elevated levels of intracellular reactive oxygen species (ROS). This situation has led to proposals that many natural stresses might be toxic specifically because they accelerate endogenous ROS formation. Such a mechanism has been convincingly demonstrated for redox-cycling compounds. However, the evidence is much weaker for most other stressors. The hypothesis that clinical antibiotics generate lethal ROS stress has attracted much attention, and the author discusses some aspects of evidence that support or oppose this idea. Importantly, even if all cellular electron flow were somehow diverted to ROS formation, the resultant doses of H2O2 and O2(-) would more likely be bacteriostatic than bacteriocidal unless key defense mechanisms were simultaneously blocked.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
-
- Kohanski MA, Dwyer DJ, Hayete B, Lawrence CA, Collins JJ. A common mechanism of cellular death induced by bactericidal antibiotics. Cell. 2007;130:797–810. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
