

Role of the unfolded protein response in regulating the mucin-dependent filamentous-growth mitogen-activated protein kinase pathway
- PMID: 25666509
- PMCID: PMC4372694
- DOI: 10.1128/MCB.01501-14
Role of the unfolded protein response in regulating the mucin-dependent filamentous-growth mitogen-activated protein kinase pathway
Abstract
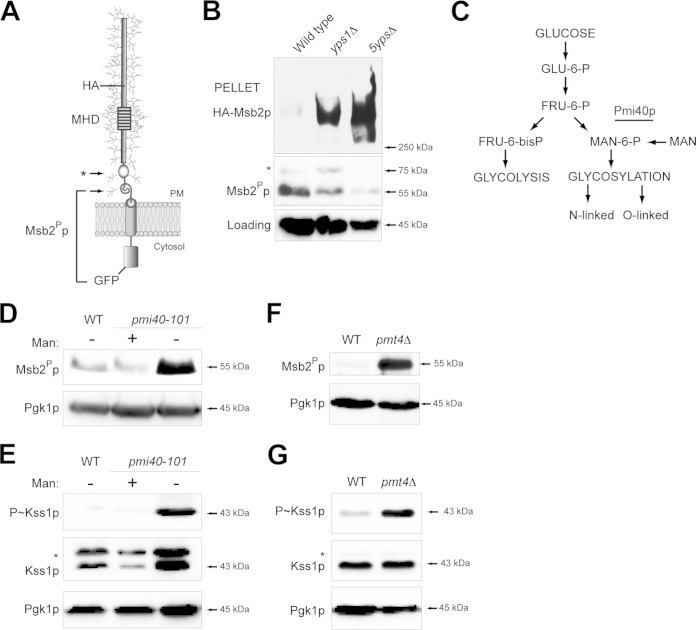
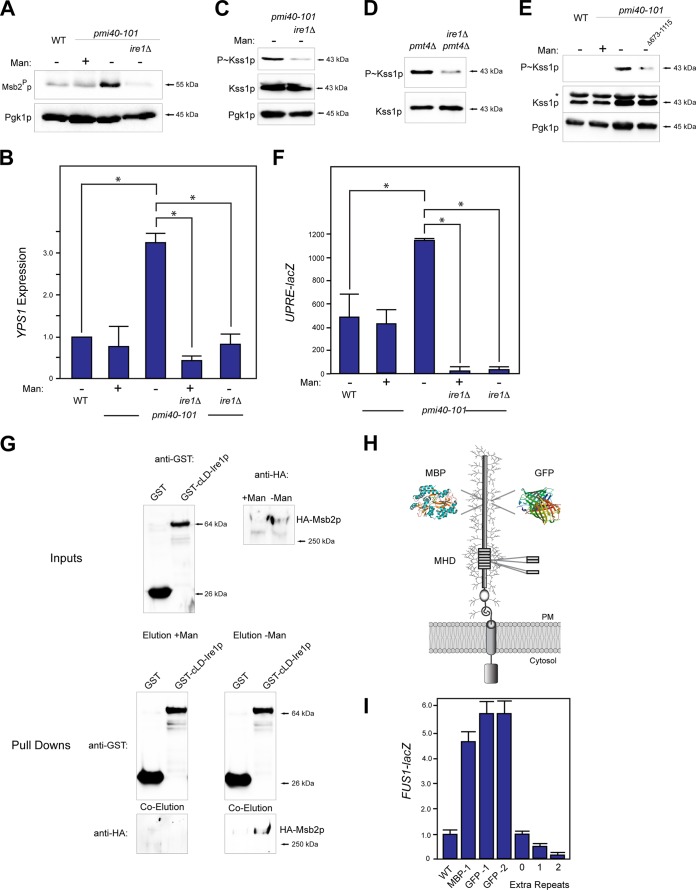
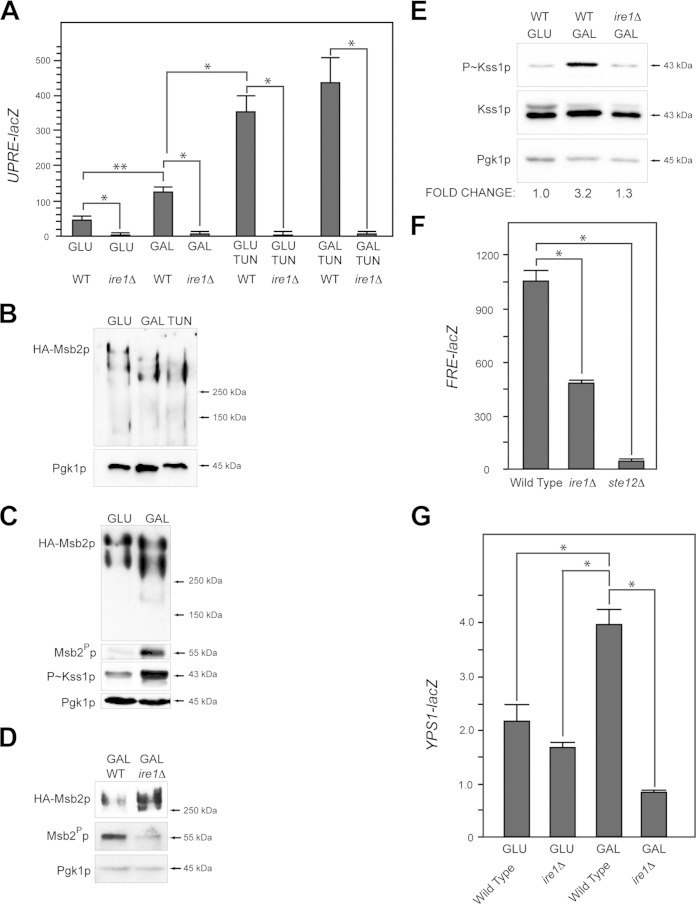
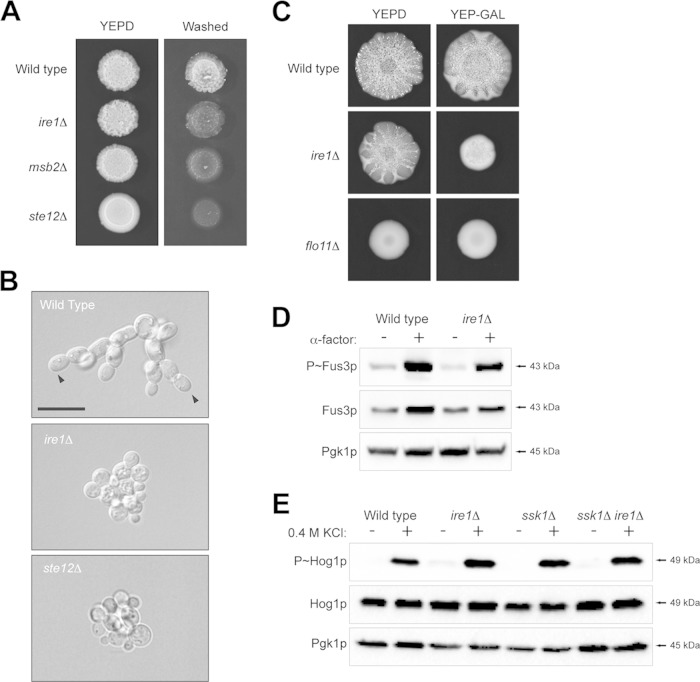
Signaling mucins are evolutionarily conserved regulators of signal transduction pathways. The signaling mucin Msb2p regulates the Cdc42p-dependent mitogen-activated protein kinase (MAPK) pathway that controls filamentous growth in yeast. The cleavage and release of the glycosylated inhibitory domain of Msb2p is required for MAPK activation. We show here that proteolytic processing of Msb2p was induced by underglycosylation of its extracellular domain. Cleavage of underglycosylated Msb2p required the unfolded protein response (UPR), a quality control (QC) pathway that operates in the endoplasmic reticulum (ER). The UPR regulator Ire1p, which detects misfolded/underglycosylated proteins in the ER, controlled Msb2p cleavage by regulating transcriptional induction of Yps1p, the major protease that processes Msb2p. Accordingly, the UPR was required for differentiation to the filamentous cell type. Cleavage of Msb2p occurred in conditional trafficking mutants that trap secretory cargo in the endomembrane system. Processed Msb2p was delivered to the plasma membrane, and its turnover by the ubiquitin ligase Rsp5p and ESCRT attenuated the filamentous-growth pathway. We speculate that the QC pathways broadly regulate signaling glycoproteins and their cognate pathways by recognizing altered glycosylation patterns that can occur in response to extrinsic cues.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
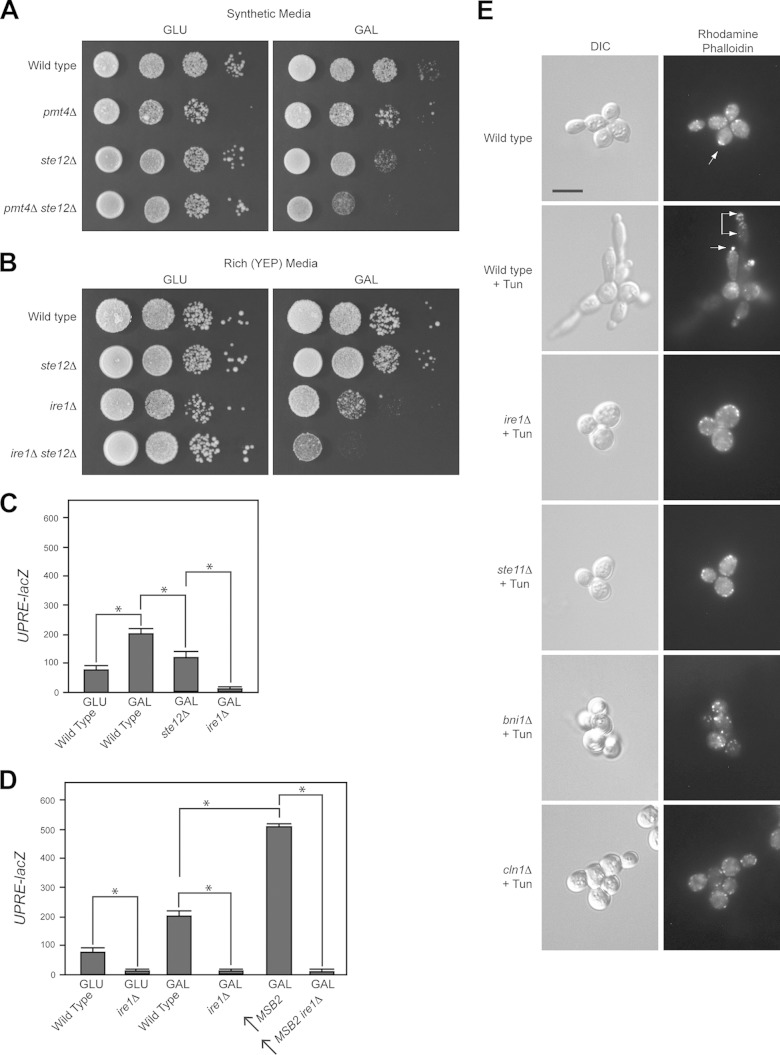
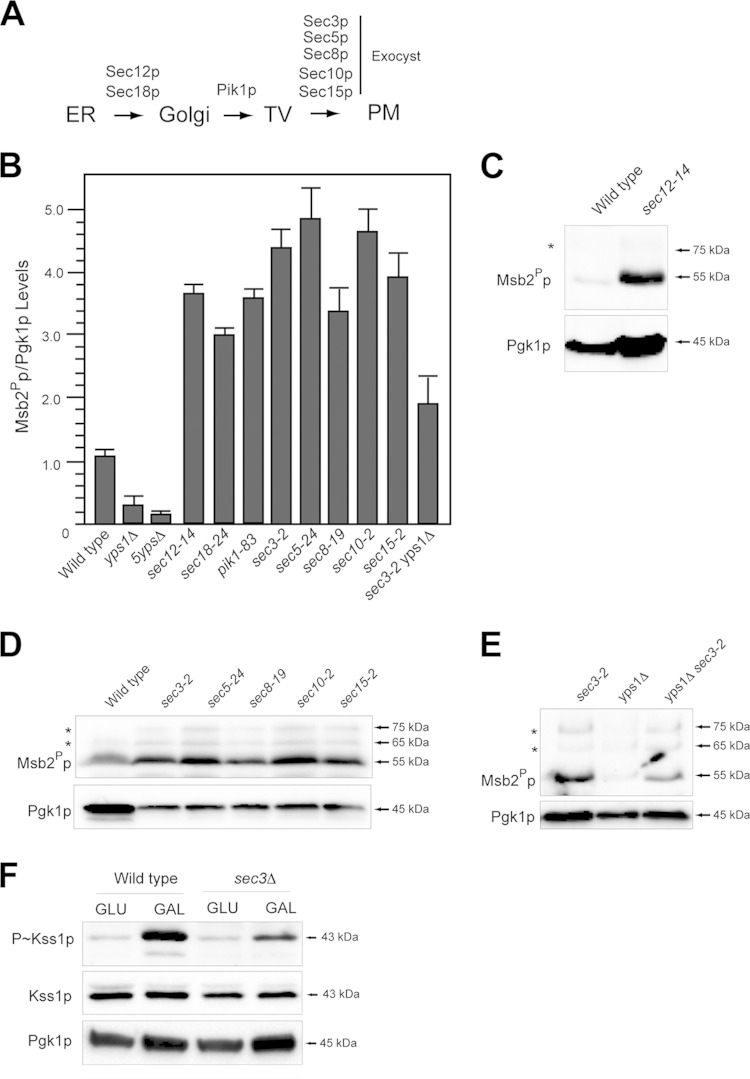
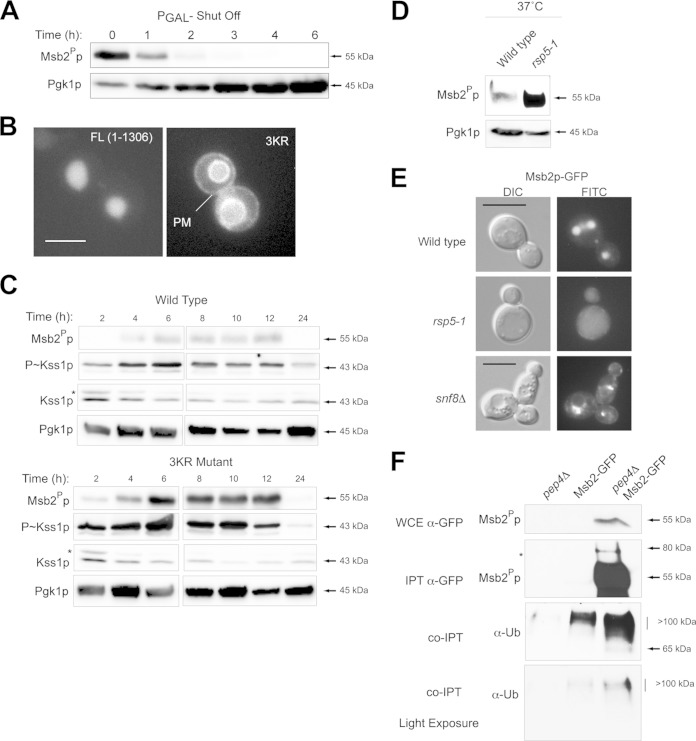
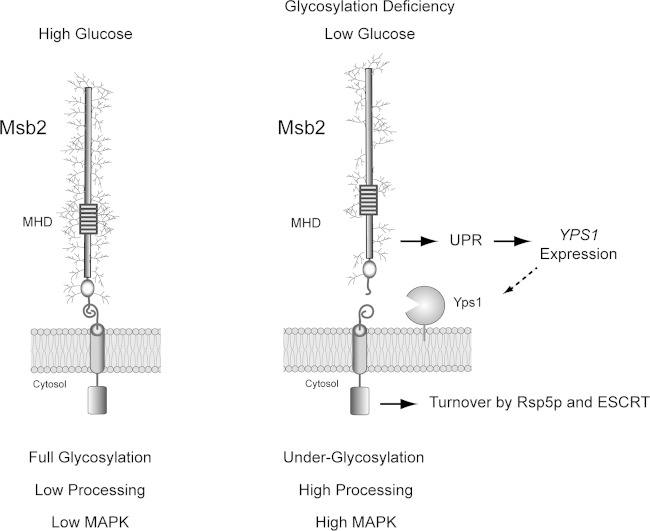
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases