

Diverse heterologous primary infections radically alter immunodominance hierarchies and clinical outcomes following H7N9 influenza challenge in mice
- PMID: 25668410
- PMCID: PMC4335497
- DOI: 10.1371/journal.ppat.1004642
Diverse heterologous primary infections radically alter immunodominance hierarchies and clinical outcomes following H7N9 influenza challenge in mice
Abstract
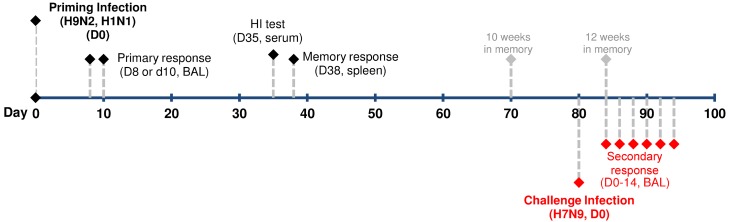
The recent emergence of a novel H7N9 influenza A virus (IAV) causing severe human infections in China raises concerns about a possible pandemic. The lack of pre-existing neutralizing antibodies in the broader population highlights the potential protective role of IAV-specific CD8(+) cytotoxic T lymphocyte (CTL) memory specific for epitopes conserved between H7N9 and previously encountered IAVs. In the present study, the heterosubtypic immunity generated by prior H9N2 or H1N1 infections significantly, but variably, reduced morbidity and mortality, pulmonary virus load and time to clearance in mice challenged with the H7N9 virus. In all cases, the recall of established CTL memory was characterized by earlier, greater airway infiltration of effectors targeting the conserved or cross-reactive H7N9 IAV peptides; though, depending on the priming IAV, each case was accompanied by distinct CTL epitope immunodominance hierarchies for the prominent K(b)PB(1703, D(b)PA(224), and D(b)NP(366) epitopes. While the presence of conserved, variable, or cross-reactive epitopes between the priming H9N2 and H1N1 and the challenge H7N9 IAVs clearly influenced any change in the immunodominance hierarchy, the changing patterns were not tied solely to epitope conservation. Furthermore, the total size of the IAV-specific memory CTL pool after priming was a better predictor of favorable outcomes than the extent of epitope conservation or secondary CTL expansion. Modifying the size of the memory CTL pool significantly altered its subsequent protective efficacy on disease severity or virus clearance, confirming the important role of heterologous priming. These findings establish that both the protective efficacy of heterosubtypic immunity and CTL immunodominance hierarchies are reflective of the immunological history of the host, a finding that has implications for understanding human CTL responses and the rational design of CTL-mediated vaccines.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Memory T cells generated by prior exposure to influenza cross react with the novel H7N9 influenza virus and confer protective heterosubtypic immunity.PLoS One. 2015 Feb 11;10(2):e0115725. doi: 10.1371/journal.pone.0115725. eCollection 2015. PLoS One. 2015. PMID: 25671696 Free PMC article.
-
Addition of a prominent epitope affects influenza A virus-specific CD8+ T cell immunodominance hierarchies when antigen is limiting.J Immunol. 2006 Sep 1;177(5):2917-25. doi: 10.4049/jimmunol.177.5.2917. J Immunol. 2006. PMID: 16920927
-
A Role of Influenza Virus Exposure History in Determining Pandemic Susceptibility and CD8+ T Cell Responses.J Virol. 2016 Jul 11;90(15):6936-6947. doi: 10.1128/JVI.00349-16. Print 2016 Aug 1. J Virol. 2016. PMID: 27226365 Free PMC article.
-
Human influenza viruses and CD8(+) T cell responses.Curr Opin Virol. 2016 Feb;16:132-142. doi: 10.1016/j.coviro.2016.01.016. Epub 2016 Mar 12. Curr Opin Virol. 2016. PMID: 26974887 Review.
-
T cell epitope engineering: an avian H7N9 influenza vaccine strategy for pandemic preparedness and response.Hum Vaccin Immunother. 2018;14(9):2203-2207. doi: 10.1080/21645515.2018.1495303. Epub 2018 Sep 5. Hum Vaccin Immunother. 2018. PMID: 30015562 Free PMC article. Review.
Cited by
-
Quantifying T Cell Cross-Reactivity: Influenza and Coronaviruses.Viruses. 2021 Sep 7;13(9):1786. doi: 10.3390/v13091786. Viruses. 2021. PMID: 34578367 Free PMC article. Review.
-
The power of memory T cells minus antibodies.Nat Immunol. 2024 Apr;25(4):594-595. doi: 10.1038/s41590-024-01796-y. Nat Immunol. 2024. PMID: 38491310 No abstract available.
-
Lung γδ T Cells Mediate Protective Responses during Neonatal Influenza Infection that Are Associated with Type 2 Immunity.Immunity. 2018 Sep 18;49(3):531-544.e6. doi: 10.1016/j.immuni.2018.07.011. Epub 2018 Aug 28. Immunity. 2018. PMID: 30170813 Free PMC article.
-
Innate and adaptive T cells in influenza disease.Front Med. 2018 Feb;12(1):34-47. doi: 10.1007/s11684-017-0606-8. Epub 2018 Jan 20. Front Med. 2018. PMID: 29352371 Review.
-
Avian Influenza Viruses, Inflammation, and CD8(+) T Cell Immunity.Front Immunol. 2016 Mar 1;7:60. doi: 10.3389/fimmu.2016.00060. eCollection 2016. Front Immunol. 2016. PMID: 26973644 Free PMC article. Review.
References
-
- Chen E, Chen Y, Fu L, Chen Z, Gong Z et al. (2013) Human infection with avian influenza A(H7N9) virus re-emerges in China in winter 2013. Euro Surveill 18 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
