

Chromatin regulation at the frontier of synthetic biology
- PMID: 25668787
- PMCID: PMC4846386
- DOI: 10.1038/nrg3900
Chromatin regulation at the frontier of synthetic biology
Abstract
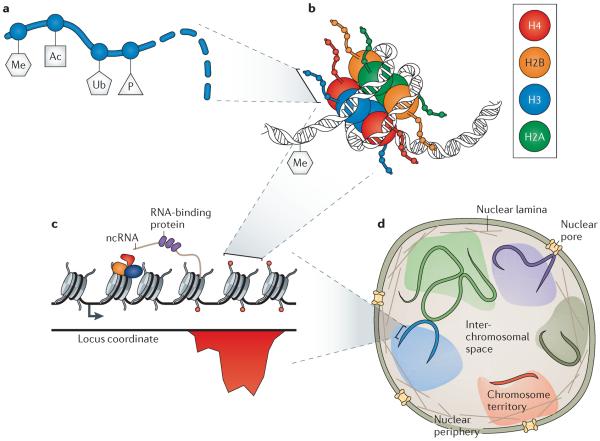
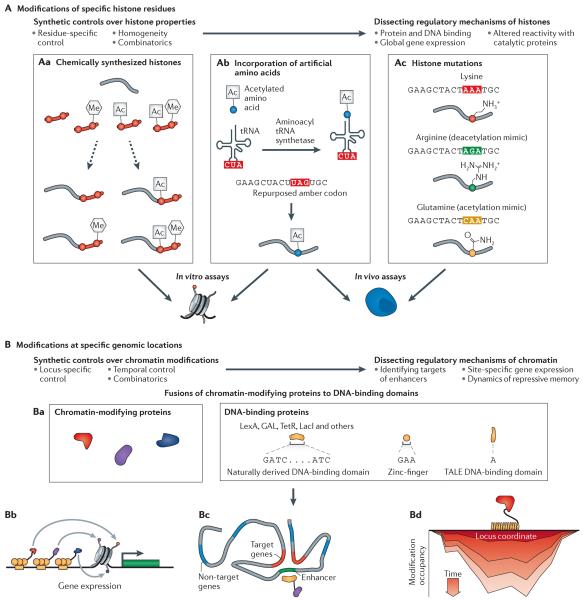
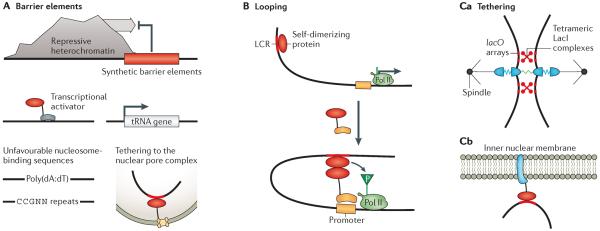
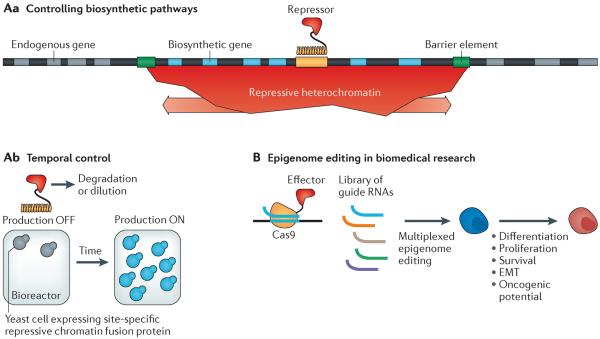
As synthetic biology approaches are extended to diverse applications throughout medicine, biotechnology and basic biological research, there is an increasing need to engineer yeast, plant and mammalian cells. Eukaryotic genomes are regulated by the diverse biochemical and biophysical states of chromatin, which brings distinct challenges, as well as opportunities, over applications in bacteria. Recent synthetic approaches, including 'epigenome editing', have allowed the direct and functional dissection of many aspects of physiological chromatin regulation. These studies lay the foundation for biomedical and biotechnological engineering applications that could take advantage of the unique combinatorial and spatiotemporal layers of chromatin regulation to create synthetic systems of unprecedented sophistication.
Figures
References
-
- Gardner TS, Cantor CR, Collins JJ. Construction of a genetic toggle switch in Escherichia coli. Nature. 2000;403:339–342. - PubMed
-
- Elowitz MB, Leibler S. A synthetic oscillatory network of transcriptional regulators. Nature. 2000;403:335–338. - PubMed
-
- Ye H, Aubel D, Fussenegger M. Synthetic mammalian gene circuits for biomedical applications. Curr. Opin. Chem. Biol. 2013;17:910–917. - PubMed
-
- Struhl K. Fundamentally different logic of gene regulation in eukaryotes and prokaryotes. Cell. 1999;98:1–4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
