

Potential effector and immunoregulatory functions of mast cells in mucosal immunity
- PMID: 25669149
- PMCID: PMC4739802
- DOI: 10.1038/mi.2014.131
Potential effector and immunoregulatory functions of mast cells in mucosal immunity
Abstract
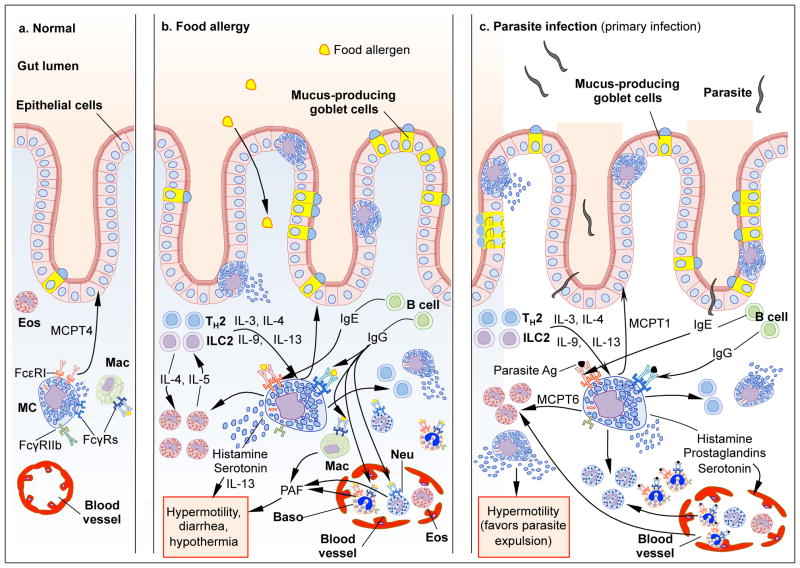
Mast cells (MCs) are cells of hematopoietic origin that normally reside in mucosal tissues, often near epithelial cells, glands, smooth muscle cells, and nerves. Best known for their contributions to pathology during IgE-associated disorders such as food allergy, asthma, and anaphylaxis, MCs are also thought to mediate IgE-associated effector functions during certain parasite infections. However, various MC populations also can be activated to express functional programs--such as secreting preformed and/or newly synthesized biologically active products--in response to encounters with products derived from diverse pathogens, other host cells (including leukocytes and structural cells), damaged tissue, or the activation of the complement or coagulation systems, as well as by signals derived from the external environment (including animal toxins, plant products, and physical agents). In this review, we will discuss evidence suggesting that MCs can perform diverse effector and immunoregulatory roles that contribute to homeostasis or pathology in mucosal tissues.
Figures

References
-
- Mekori YA, Metcalfe DD. Mast cells in innate immunity. Immunol Rev. 2000;173:131–140. - PubMed
-
- Metz M, Grimbaldeston MA, Nakae S, Piliponsky AM, Tsai M, Galli SJ. Mast cells in the promotion and limitation of chronic inflammation. Immunol Rev. 2007;217:304–328. - PubMed
-
- Moon TC, St Laurent CD, Morris KE, Marcet C, Yoshimura T, Sekar Y, et al. Advances in mast cell biology: new understanding of heterogeneity and function. Mucosal Immunol. 2010;3(2):111–128. - PubMed
-
- Gurish MF, Austen KF. Developmental origin and functional specialization of mast cell subsets. Immunity. 2012;37(1):25–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
