

Quantitative analysis of acetyl-CoA production in hypoxic cancer cells reveals substantial contribution from acetate
- PMID: 25671109
- PMCID: PMC4322440
- DOI: 10.1186/2049-3002-2-23
Quantitative analysis of acetyl-CoA production in hypoxic cancer cells reveals substantial contribution from acetate
Abstract
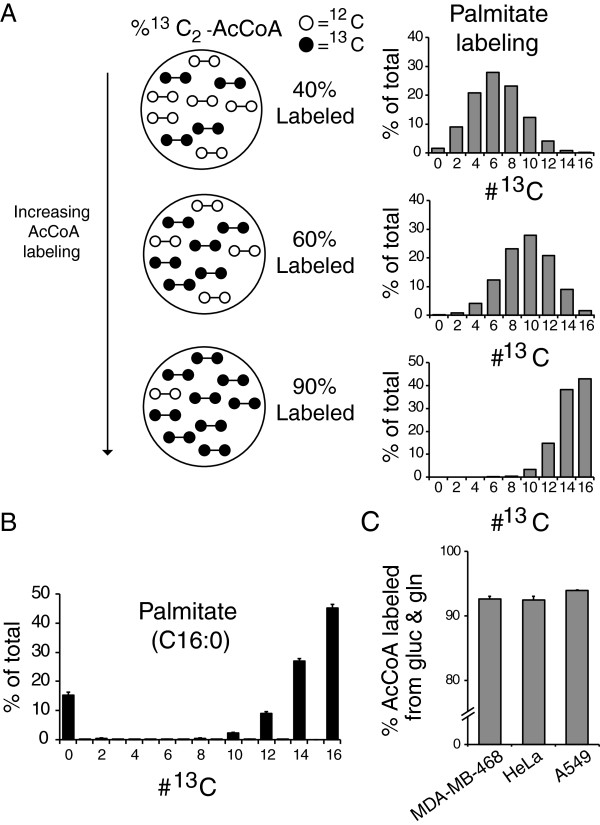
Background: Cell growth requires fatty acids for membrane synthesis. Fatty acids are assembled from 2-carbon units in the form of acetyl-CoA (AcCoA). In nutrient and oxygen replete conditions, acetyl-CoA is predominantly derived from glucose. In hypoxia, however, flux from glucose to acetyl-CoA decreases, and the fractional contribution of glutamine to acetyl-CoA increases. The significance of other acetyl-CoA sources, however, has not been rigorously evaluated. Here we investigate quantitatively, using (13)C-tracers and mass spectrometry, the sources of acetyl-CoA in hypoxia.
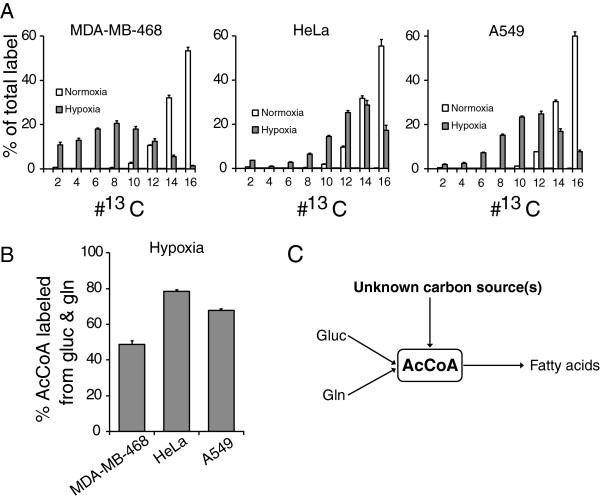
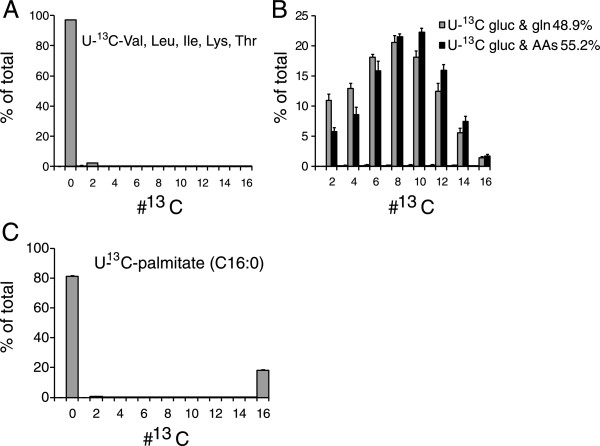
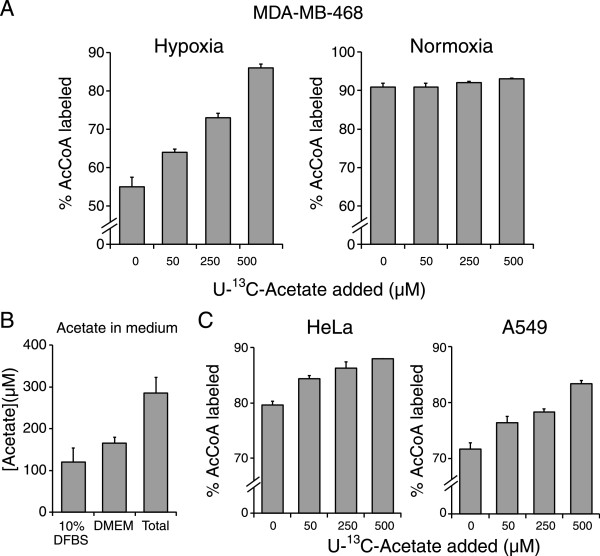
Results: In normoxic conditions, cultured cells produced more than 90% of acetyl-CoA from glucose and glutamine-derived carbon. In hypoxic cells, this contribution dropped, ranging across cell lines from 50% to 80%. Thus, under hypoxia, one or more additional substrates significantly contribute to acetyl-CoA production. (13)C-tracer experiments revealed that neither amino acids nor fatty acids are the primary source of this acetyl-CoA. Instead, the main additional source is acetate. A large contribution from acetate occurs despite it being present in the medium at a low concentration (50-500 μM).
Conclusions: Acetate is an important source of acetyl-CoA in hypoxia. Inhibition of acetate metabolism may impair tumor growth.
Keywords: 13C-tracing; Acetate; Acetyl-CoA; Cancer metabolism; Fatty acids; Hypoxia; Lipogenesis; Mass spectrometry; Palmitate.
Figures
Similar articles
-
Metabolic engineering for efficient supply of acetyl-CoA from different carbon sources in Escherichia coli.Microb Cell Fact. 2019 Aug 6;18(1):130. doi: 10.1186/s12934-019-1177-y. Microb Cell Fact. 2019. PMID: 31387584 Free PMC article.
-
Metabolic and mind shifts: from glucose to glutamine and acetate addictions in cancer.Curr Opin Clin Nutr Metab Care. 2015 Jul;18(4):346-53. doi: 10.1097/MCO.0000000000000178. Curr Opin Clin Nutr Metab Care. 2015. PMID: 26001655 Review.
-
Effects of dichloroacetate on the metabolism of glucose, pyruvate, acetate, 3-hydroxybutyrate and palmitate in rat diaphragm and heart muscle in vitro and on extraction of glucose, lactate, pyruvate and free fatty acids by dog heart in vivo.Biochem J. 1973 Aug;134(4):1067-81. doi: 10.1042/bj1341067. Biochem J. 1973. PMID: 4762752 Free PMC article.
-
Probing in vivo metabolism by stable isotope labeling of storage lipids and proteins in developing Brassica napus embryos.Plant Physiol. 2002 Sep;130(1):347-61. doi: 10.1104/pp.004275. Plant Physiol. 2002. PMID: 12226514 Free PMC article.
-
Acetate/acetyl-CoA metabolism associated with cancer fatty acid synthesis: overview and application.Cancer Lett. 2015 Jan 28;356(2 Pt A):211-6. doi: 10.1016/j.canlet.2014.02.019. Epub 2014 Feb 22. Cancer Lett. 2015. PMID: 24569091 Review.
Cited by
-
The combination of exon sequencing and metabolomics to establish a molecular typing system for gastric cancer.Heliyon. 2024 Jul 9;10(15):e34317. doi: 10.1016/j.heliyon.2024.e34317. eCollection 2024 Aug 15. Heliyon. 2024. PMID: 39170180 Free PMC article.
-
Inferring mitochondrial and cytosolic metabolism by coupling isotope tracing and deconvolution.Nat Commun. 2023 Nov 18;14(1):7525. doi: 10.1038/s41467-023-42824-z. Nat Commun. 2023. PMID: 37980339 Free PMC article.
-
Metabolic Regulation of Lysine Acetylation: Implications in Cancer.Subcell Biochem. 2022;100:393-426. doi: 10.1007/978-3-031-07634-3_12. Subcell Biochem. 2022. PMID: 36301501 Review.
-
A Nutrient-Based Cellular Model to Characterize Acetylation-Dependent Protein-Protein Interactions.Front Mol Biosci. 2022 Mar 23;9:831758. doi: 10.3389/fmolb.2022.831758. eCollection 2022. Front Mol Biosci. 2022. PMID: 35402505 Free PMC article.
-
Acetyl-CoA Synthetase 2 as a Therapeutic Target in Tumor Metabolism.Cancers (Basel). 2022 Jun 12;14(12):2896. doi: 10.3390/cancers14122896. Cancers (Basel). 2022. PMID: 35740562 Free PMC article. Review.
References
-
- Vander Heiden MG, Lunt SY, Dayton TL, Fiske BP, Israelsen WJ, Mattaini KR, Vokes NI, Stephanopoulos G, Cantley LC, Metallo CM, Locasale JW. Metabolic pathway alterations that support cell proliferation. Cold Spring Harb Symp Quant Biol. 2011;76:325–334. doi: 10.1101/sqb.2012.76.010900. - DOI - PubMed
-
- Luyimbazi D, Akcakanat A, McAuliffe PF, Zhang L, Singh G, Gonzalez-Angulo AM, Chen H, Do K-A, Zheng Y, Hung MC, Mills GB, Meric-Bernstam F. Rapamycin regulates stearoyl CoA desaturase 1 expression in breast cancer. Mol Cancer Ther. 2010;9:2770–2784. doi: 10.1158/1535-7163.MCT-09-0980. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
