

Mitochondrial and cytoplasmic ROS have opposing effects on lifespan
- PMID: 25671321
- PMCID: PMC4335496
- DOI: 10.1371/journal.pgen.1004972
Mitochondrial and cytoplasmic ROS have opposing effects on lifespan
Abstract
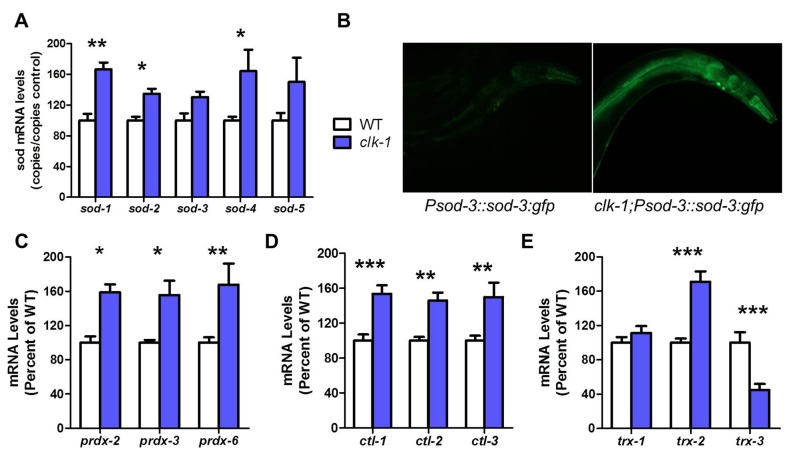
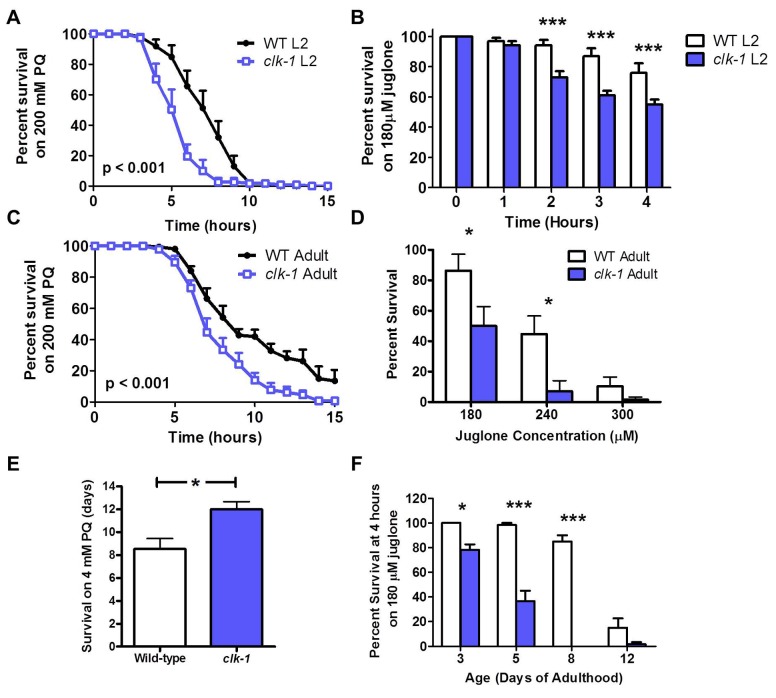
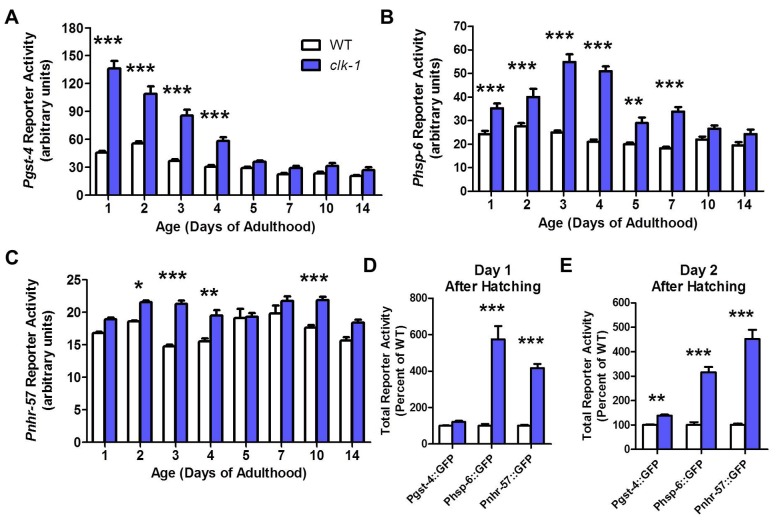
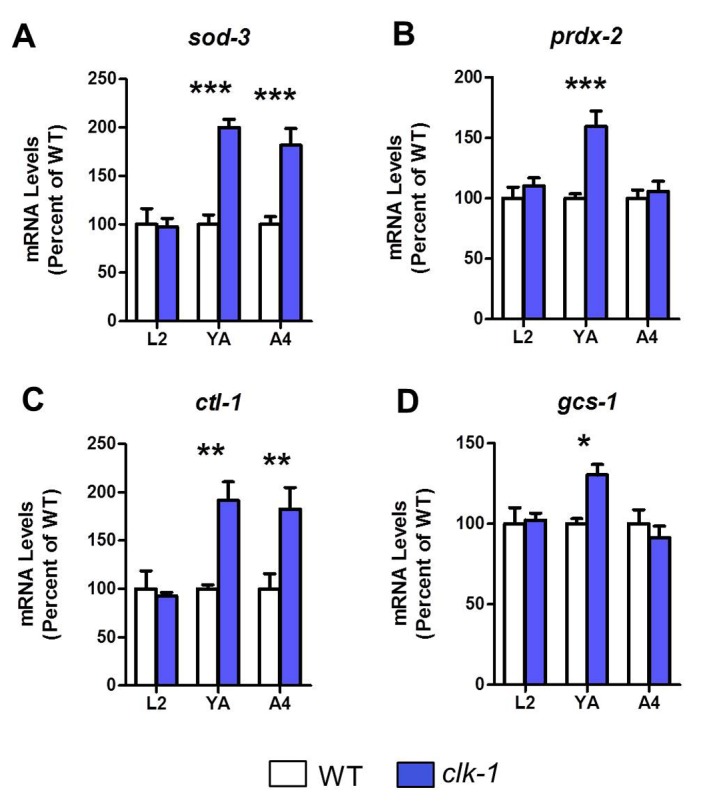
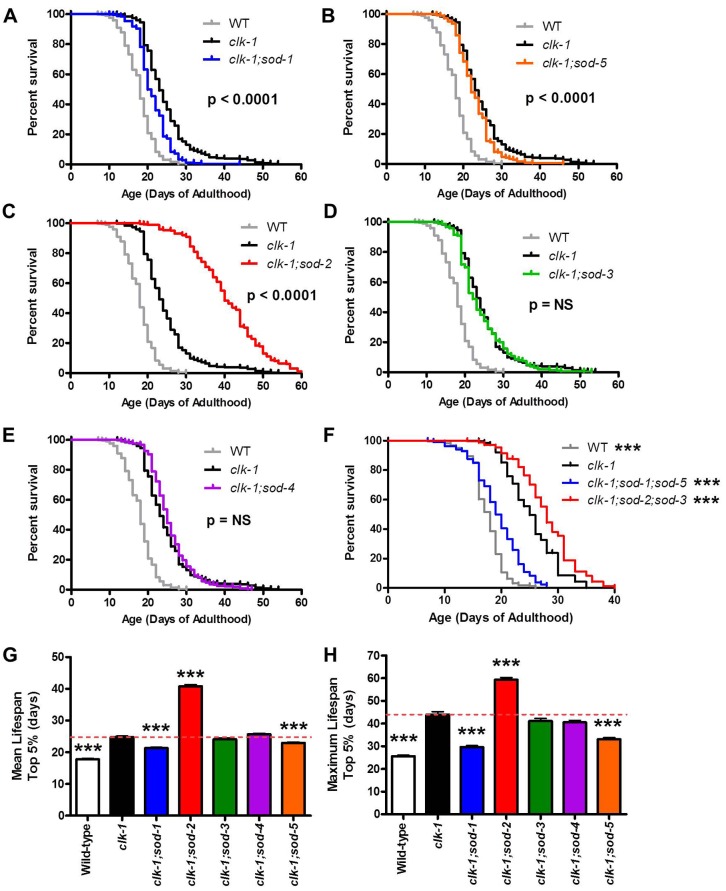
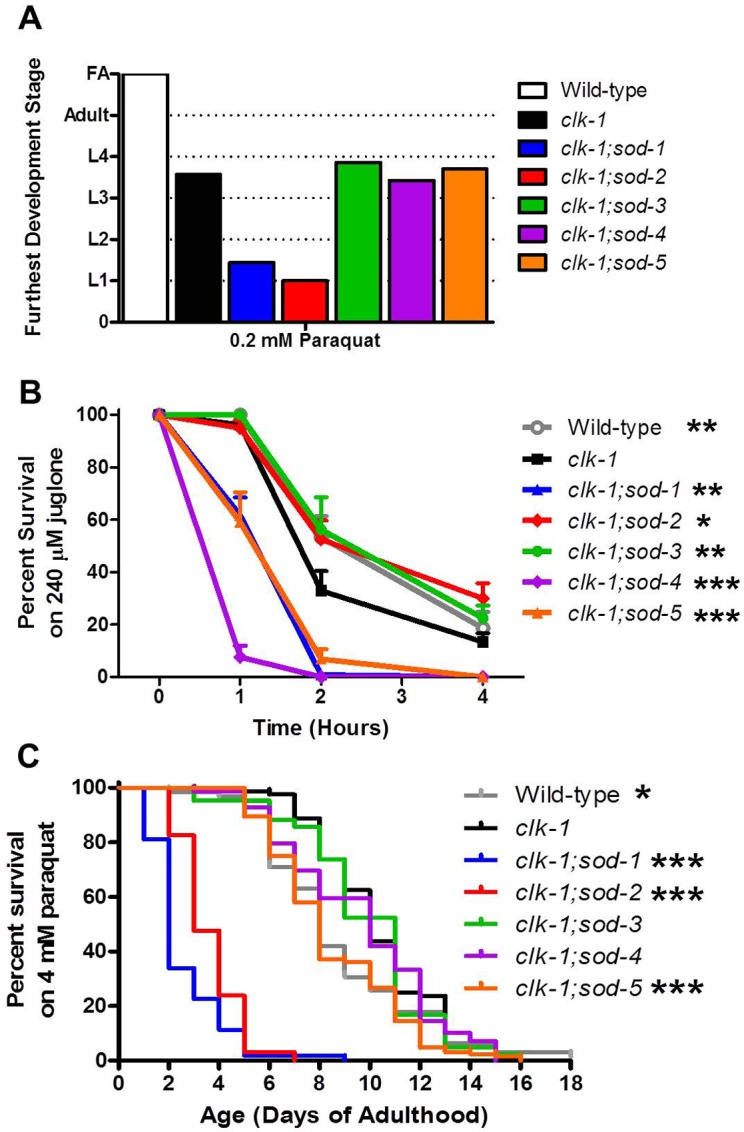
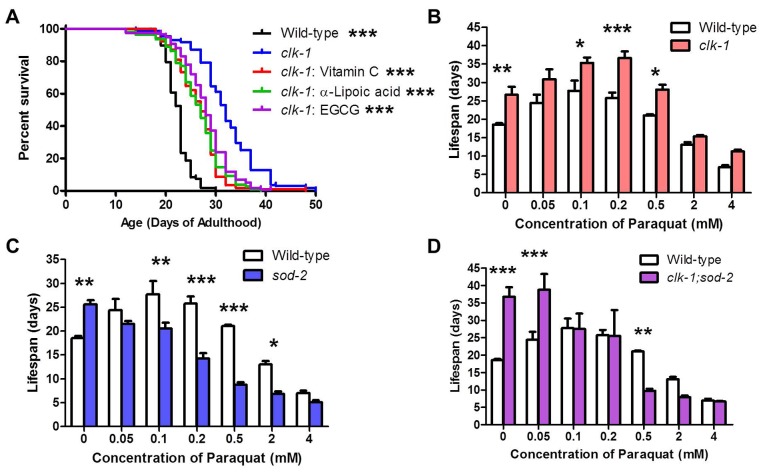
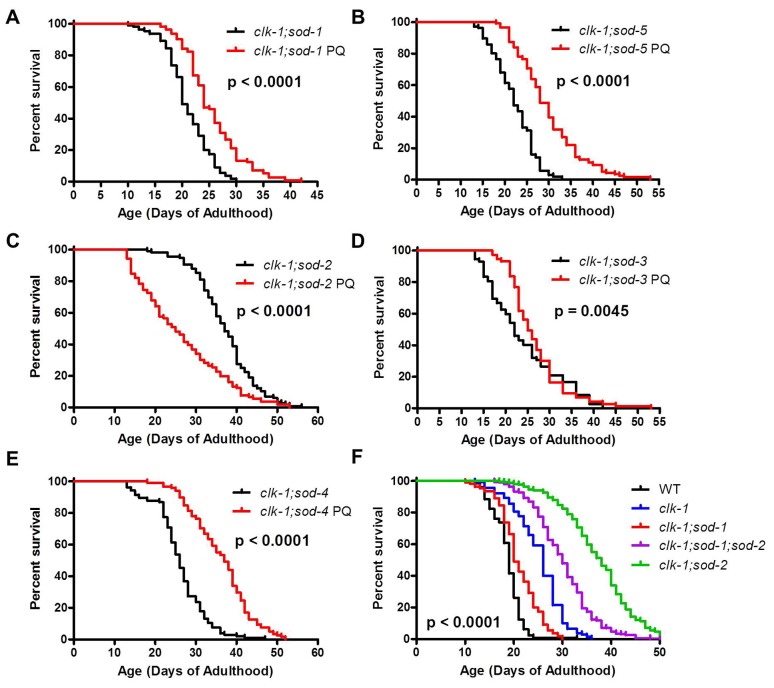
Reactive oxygen species (ROS) are highly reactive, oxygen-containing molecules that can cause molecular damage within the cell. While the accumulation of ROS-mediated damage is widely believed to be one of the main causes of aging, ROS also act in signaling pathways. Recent work has demonstrated that increasing levels of superoxide, one form of ROS, through treatment with paraquat, results in increased lifespan. Interestingly, treatment with paraquat robustly increases the already long lifespan of the clk-1 mitochondrial mutant, but not other long-lived mitochondrial mutants such as isp-1 or nuo-6. To genetically dissect the subcellular compartment in which elevated ROS act to increase lifespan, we deleted individual superoxide dismutase (sod) genes in clk-1 mutants, which are sensitized to ROS. We find that only deletion of the primary mitochondrial sod gene, sod-2 results in increased lifespan in clk-1 worms. In contrast, deletion of either of the two cytoplasmic sod genes, sod-1 or sod-5, significantly decreases the lifespan of clk-1 worms. Further, we show that increasing mitochondrial superoxide levels through deletion of sod-2 or treatment with paraquat can still increase lifespan in clk-1;sod-1 double mutants, which live shorter than clk-1 worms. The fact that mitochondrial superoxide can increase lifespan in worms with a detrimental level of cytoplasmic superoxide demonstrates that ROS have a compartment specific effect on lifespan - elevated ROS in the mitochondria acts to increase lifespan, while elevated ROS in the cytoplasm decreases lifespan. This work also suggests that both ROS-dependent and ROS-independent mechanisms contribute to the longevity of clk-1 worms.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
