

Methamphetamine accelerates cellular senescence through stimulation of de novo ceramide biosynthesis
- PMID: 25671639
- PMCID: PMC4324822
- DOI: 10.1371/journal.pone.0116961
Methamphetamine accelerates cellular senescence through stimulation of de novo ceramide biosynthesis
Abstract
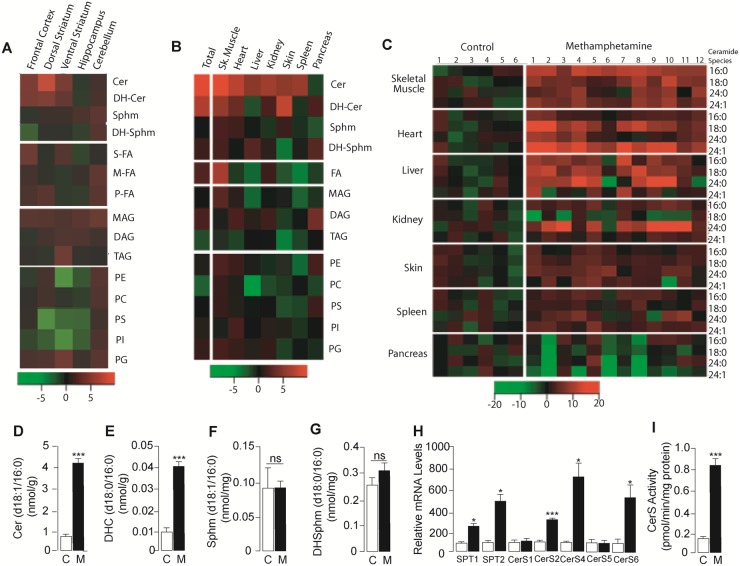
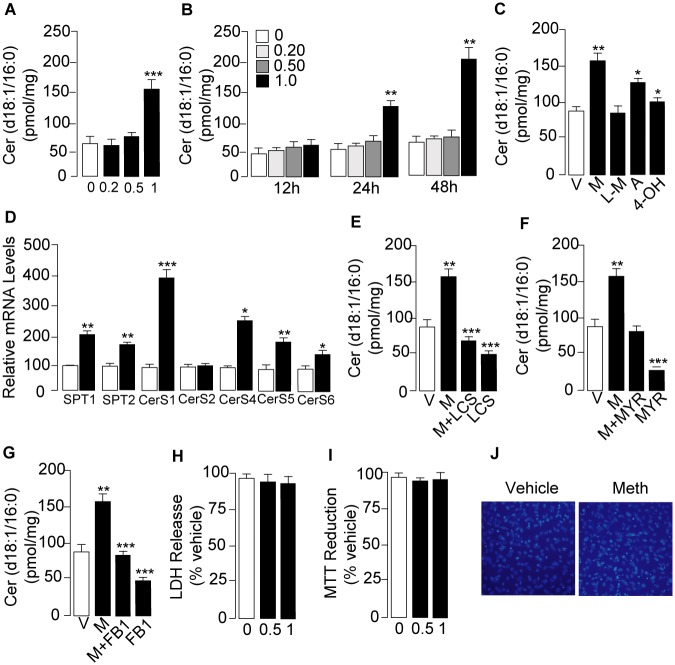
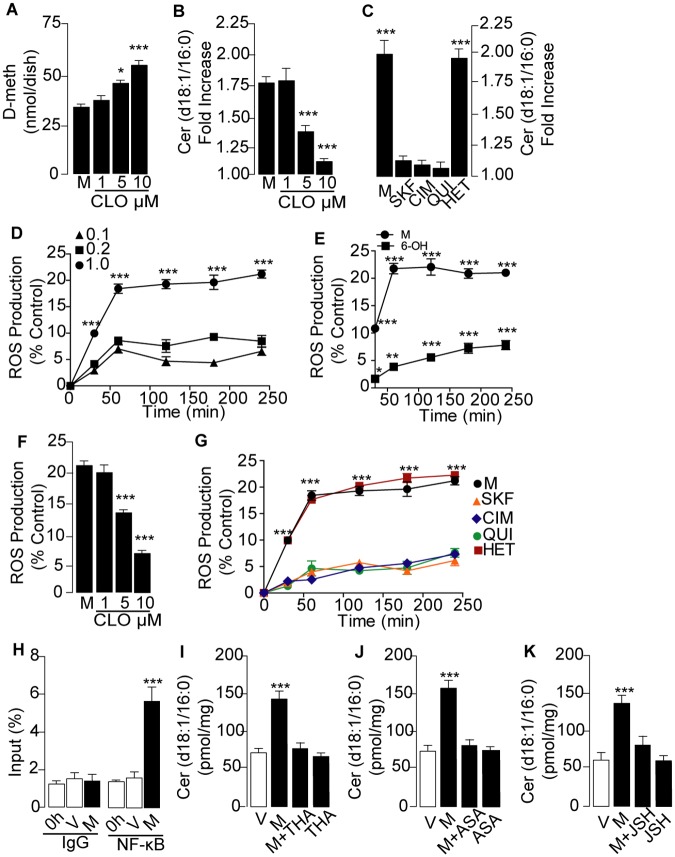
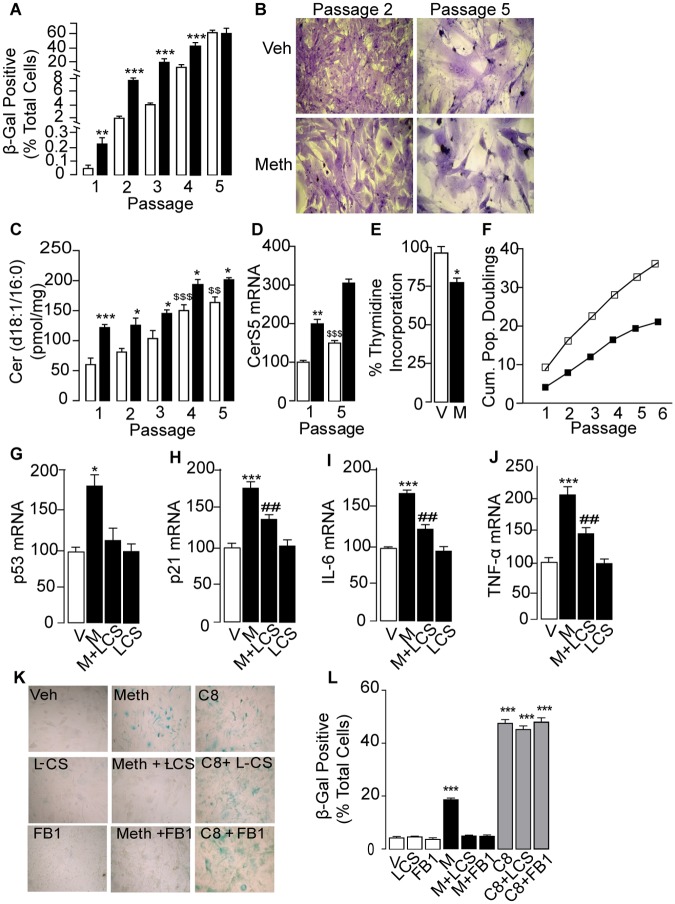
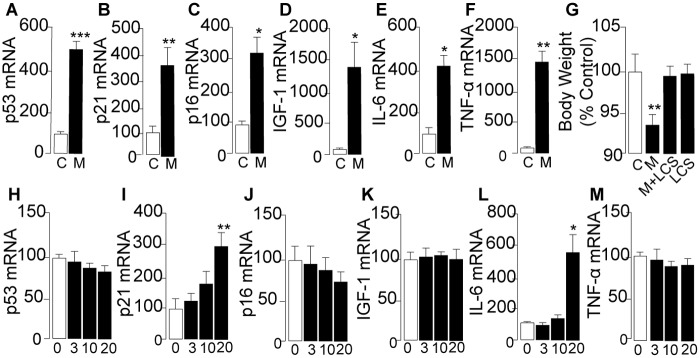
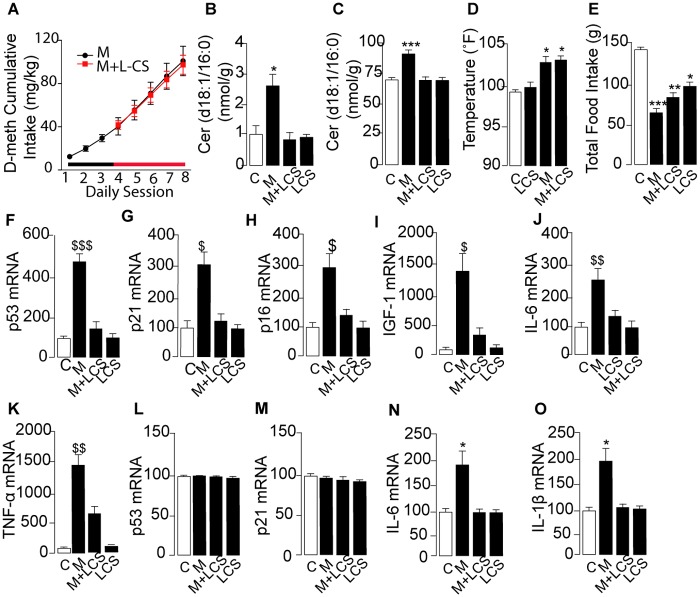
Methamphetamine is a highly addictive psychostimulant that causes profound damage to the brain and other body organs. Post mortem studies of human tissues have linked the use of this drug to diseases associated with aging, such as coronary atherosclerosis and pulmonary fibrosis, but the molecular mechanism underlying these findings remains unknown. Here we used functional lipidomics and transcriptomics experiments to study abnormalities in lipid metabolism in select regions of the brain and, to a greater extent, peripheral organs and tissues of rats that self-administered methamphetamine. Experiments in various cellular models (primary mouse fibroblasts and myotubes) allowed us to investigate the molecular mechanisms of systemic inflammation and cellular aging related to methamphetamine abuse. We report now that methamphetamine accelerates cellular senescence and activates transcription of genes involved in cell-cycle control and inflammation by stimulating production of the sphingolipid messenger ceramide. This pathogenic cascade is triggered by reactive oxygen species, likely generated through methamphetamine metabolism via cytochrome P450, and involves the recruitment of nuclear factor-κB (NF-κB) to induce expression of enzymes in the de novo pathway of ceramide biosynthesis. Inhibitors of NF-κB signaling and ceramide formation prevent methamphetamine-induced senescence and systemic inflammation in rats self-administering the drug, attenuating their health deterioration. The results suggest new therapeutic strategies to reduce the adverse consequences of methamphetamine abuse and improve effectiveness of abstinence treatments.
Conflict of interest statement
Figures
Similar articles
-
Asiatic acid attenuates methamphetamine-induced neuroinflammation and neurotoxicity through blocking of NF-kB/STAT3/ERK and mitochondria-mediated apoptosis pathway.J Neuroinflammation. 2017 Dec 11;14(1):240. doi: 10.1186/s12974-017-1009-0. J Neuroinflammation. 2017. PMID: 29228978 Free PMC article.
-
Methamphetamine-induced TNF-alpha gene expression and activation of AP-1 in discrete regions of mouse brain: potential role of reactive oxygen intermediates and lipid peroxidation.Neuromolecular Med. 2002;2(1):71-85. doi: 10.1385/NMM:2:1:71. Neuromolecular Med. 2002. PMID: 12230306
-
Extended-access methamphetamine self-administration elicits neuroinflammatory response along with blood-brain barrier breakdown.Brain Behav Immun. 2017 May;62:306-317. doi: 10.1016/j.bbi.2017.02.017. Epub 2017 Feb 24. Brain Behav Immun. 2017. PMID: 28237710
-
Emerging role of NF-κB signaling in the induction of senescence-associated secretory phenotype (SASP).Cell Signal. 2012 Apr;24(4):835-45. doi: 10.1016/j.cellsig.2011.12.006. Epub 2011 Dec 11. Cell Signal. 2012. PMID: 22182507 Review.
-
[Underlying Mechanisms of Methamphetamine-Induced Self-Injurious Behavior and Lethal Effects in Mice].Nihon Eiseigaku Zasshi. 2018;73(1):51-56. doi: 10.1265/jjh.73.51. Nihon Eiseigaku Zasshi. 2018. PMID: 29386447 Review. Japanese.
Cited by
-
Integrative Analysis of Global Gene Expression Identifies Opposite Patterns of Reactive Astrogliosis in Aged Human Prefrontal Cortex.Brain Sci. 2018 Dec 19;8(12):227. doi: 10.3390/brainsci8120227. Brain Sci. 2018. PMID: 30572619 Free PMC article.
-
Dopaminergic Neuron-Specific Deletion of p53 Gene Attenuates Methamphetamine Neurotoxicity.Neurotox Res. 2017 Aug;32(2):218-230. doi: 10.1007/s12640-017-9723-z. Epub 2017 Mar 24. Neurotox Res. 2017. PMID: 28342134 Free PMC article.
-
Revealing Metabolic Perturbation Following Heavy Methamphetamine Abuse by Human Hair Metabolomics and Network Analysis.Int J Mol Sci. 2020 Aug 21;21(17):6041. doi: 10.3390/ijms21176041. Int J Mol Sci. 2020. PMID: 32839415 Free PMC article.
-
Early onset cardiovascular disease related to methamphetamine use is most striking in individuals under 30: A retrospective chart review.Addict Behav Rep. 2022 May 18;15:100435. doi: 10.1016/j.abrep.2022.100435. eCollection 2022 Jun. Addict Behav Rep. 2022. PMID: 35620216 Free PMC article.
-
The identification of small molecule inhibitors of the plant inositol phosphorylceramide synthase which demonstrate herbicidal activity.Sci Rep. 2019 May 30;9(1):8083. doi: 10.1038/s41598-019-44544-1. Sci Rep. 2019. PMID: 31147620 Free PMC article.
References
-
- Fleckenstein AE, Volz TJ, Riddle EL, Gibb JW, Hanson GR (2007) New insights into the mechanism of action of amphetamines. Annu Rev Pharmacol Toxicol 47: 681–698. - PubMed
-
- Volkow ND, Chang L, Wang GJ, Fowler JS, Ding YS, et al. (2001) Low level of brain dopamine D2 receptors in methamphetamine abusers: association with metabolism in the orbitofrontal cortex. Am J Psychiatry 158: 2015–2021. - PubMed
-
- Wang GJ, Volkow ND, Thanos PK, Fowler JS (2004) Similarity between obesity and drug addiction as assessed by neurofunctional imaging: a concept review. J Addict Dis 23: 39–53. - PubMed
-
- Acikgoz O, Gonenc S, Kayatekin BM, Uysal N, Pekcetin C, et al. (1998) Methamphetamine causes lipid peroxidation and an increase in superoxide dismutase activity in the rat striatum. Brain Res 813: 200–202. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
