

Regulation of mitochondrial pyruvate uptake by alternative pyruvate carrier complexes
- PMID: 25672363
- PMCID: PMC4388599
- DOI: 10.15252/embj.201490197
Regulation of mitochondrial pyruvate uptake by alternative pyruvate carrier complexes
Abstract
At the pyruvate branch point, the fermentative and oxidative metabolic routes diverge. Pyruvate can be transformed either into lactate in mammalian cells or into ethanol in yeast, or transported into mitochondria to fuel ATP production by oxidative phosphorylation. The recently discovered mitochondrial pyruvate carrier (MPC), encoded by MPC1, MPC2, and MPC3 in yeast, is required for uptake of pyruvate into the organelle. Here, we show that while expression of Mpc1 is not dependent on the carbon source, expression of Mpc2 and Mpc3 is specific to fermentative or respiratory conditions, respectively. This gives rise to two alternative carrier complexes that we have termed MPCFERM and MPCOX. By constitutively expressing the two alternative complexes in yeast deleted for all three endogenous genes, we show that MPCOX has a higher transport activity than MPCFERM, which is dependent on the C-terminus of Mpc3. We propose that the alternative MPC subunit expression in yeast provides a way of adapting cellular metabolism to the nutrient availability.
Keywords: metabolism; mitochondria; mitochondrial pyruvate carrier; pyruvate branch point.
© 2015 The Authors.
Figures
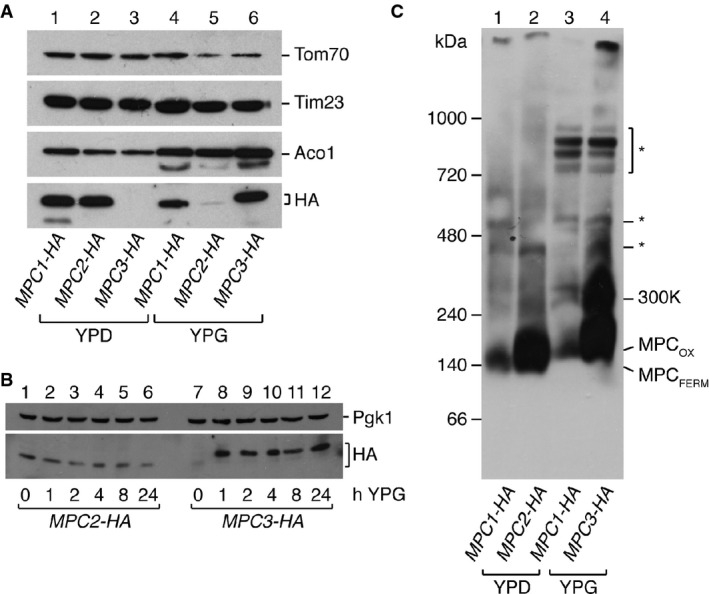
Mpc1 and Mpc2 are present under fermentative conditions, while Mpc1 and Mpc3 are present under respiratory conditions. MPC subunits were detected by Western blot in mitochondria isolated from yeast cells grown in glucose (YPD) or glycerol (YPG). Mitochondrial matrix protein aconitase (Aco1), 23-kDa translocase of the inner membrane subunit (Tim23), and 70-kDa translocase of the outer membrane subunit (Tom70) were used as loading controls.
Kinetics of the switch from Mpc2 to Mpc3. Yeast cells were grown in YPD until mid-log phase and then shifted to YPG medium. Whole cell extracts were taken at the indicated time points after the shift, and HA-tagged Mpc2 and Mpc3 detected by Western blotting. Mpc3 is induced very rapidly by switch from YPD to YPG, while Mpc2 levels decrease only gradually.
Alternative pyruvate carrier complexes MPCFERM and MPCOX are detected by BN-PAGE. Isolated mitochondria expressing HA-tagged MPC proteins were run on blue native gels, and MPC complexes detected with an HA-specific antibody. MPCFERM consisting of Mpc1 and Mpc2 is present under fermentative conditions, and MPCOX consisting of Mpc1 and Mpc3 is present under respiratory conditions. In addition, a complex of ˜300 kDa is observed under respiratory conditions that contains Mpc1 and Mpc3. Non-specific reactivity of the anti-HA antibody toward respiratory chain complexes that are highly abundant in oxidative growth conditions is denoted by an asterisk (*).
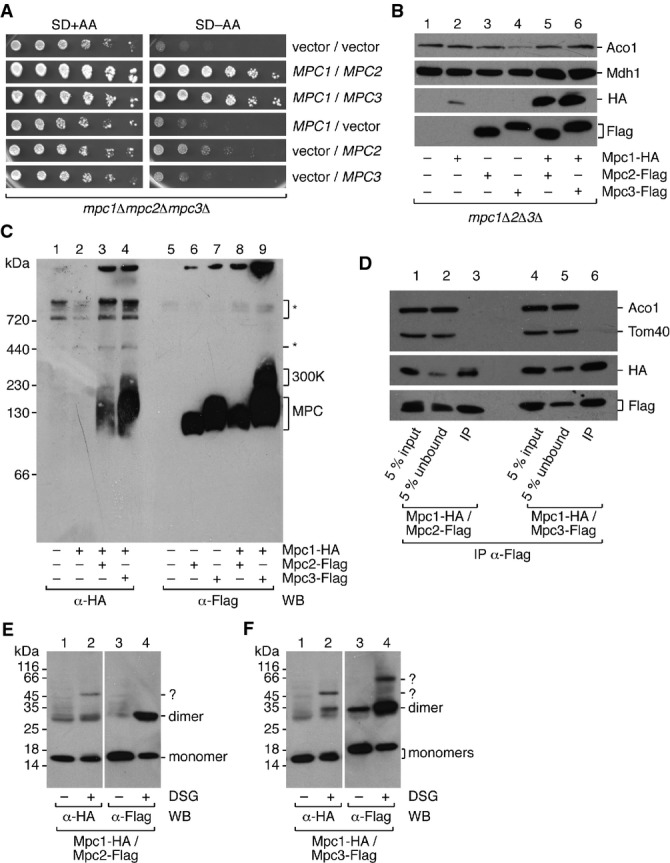
A Constitutive expression of either MPCFERM or MPCOX can rescue the slow growth phenotype of the mpc1Δmpc2Δmpc3Δ triple deletion mutant. Growth tests were performed with cells transformed with MPC1, MPC2, or MPC3 expression plasmids or with empty vectors in indicated combinations. A serial dilution of yeast cells was spotted on agar plates with glucose-containing synthetic minimal medium with (SD+AA) or without (SD−AA) amino acids.
B Plasmid-encoded MPC proteins can be detected in isolated mitochondria when Mpc1 is co-expressed with either Mpc2 or Mpc3. Cells were grown in selective medium containing glycerol as a carbon source. MPC subunits were constitutively expressed from plasmids in indicated combinations in the mpc1Δmpc2Δmpc3Δ background and detected by Western blot using anti-HA or anti-Flag antibodies. Mitochondrial enzymes aconitase (Aco1) and malate dehydrogenase (Mdh1) were used as loading controls.
C BN-PAGE confirms the formation of MPCFERM and MPCOX complexes after constitutive plasmid-based protein expression. Non-specific reactivity of the antibody is denoted by an asterisk (*). It has to be noted that all complexes migrate slightly faster than in Fig1C, most likely because MPC proteins are tagged with a single epitope tag as opposed to the 3HA-tag in Fig1C.
D Mpc1 is co-immunoprecipitated with Mpc2 or Mpc3. Mitochondria isolated from cells harboring appropriate expression plasmids and grown in glycerol-containing medium were lysed in 1% digitonin, and either Mpc2-Flag or Mpc3-Flag was immunoprecipitated. Mpc1-HA was detected by Western blot with an antibody directed against the epitope tag. The matrix protein aconitase (Aco1) and 40-kDa translocase of the outer membrane subunit (Tom40) were not co-immunoprecipitated.
E, F Stoichiometry of MPC complexes. Chemical cross-linking indicates that Mpc1/Mpc2 and Mpc1/Mpc3 form heterodimers of the predicted molecular weight. DSG, disuccimidyl glutarate. Cross-linking products with unknown partners are labeled with a question mark (?).
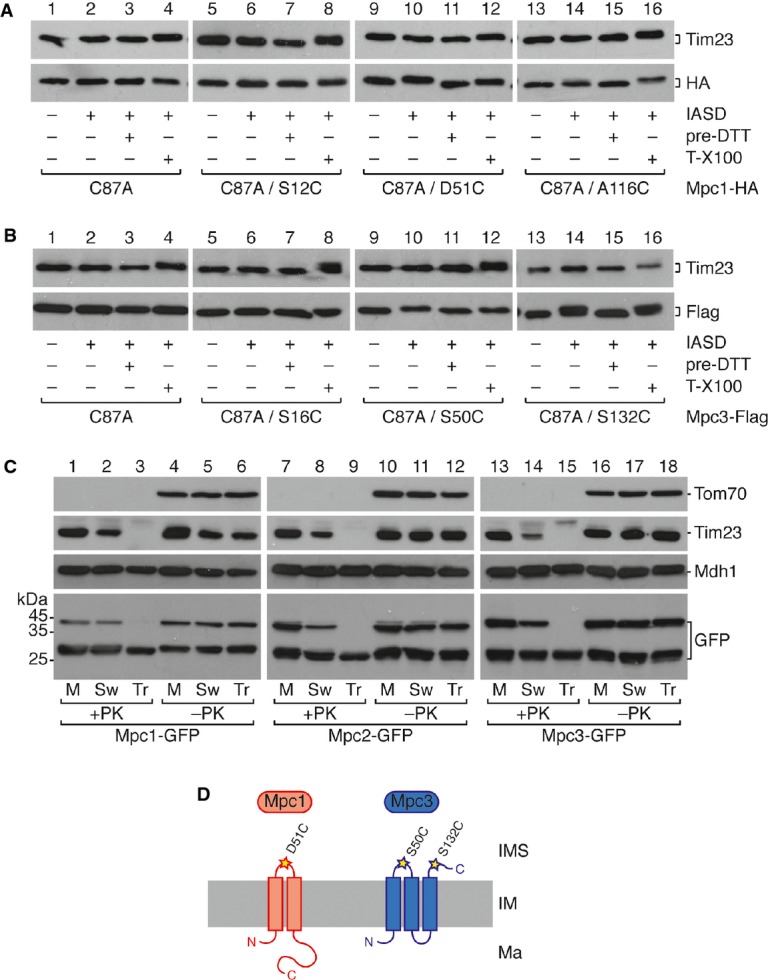
A, B Determination of MPC membrane topology by IASD labeling of isolated mitochondria expressing the indicated single cysteine variants of Mpc1 (A) and Mpc3 (B). Mitochondria were treated with IASD as indicated. Negative [quenching by pretreatment with DTT (pre-DTT)] and positive [lysis with 0.5% Triton X-100 (T-X100)] controls for the labeling reaction are shown. As a further control, also translocase of the inner membrane subunit Tim23 is detected by Western blot, which contains three endogenous cysteines, all of which are located in membrane-embedded regions. Labeling of an IMS-exposed cysteine with IASD is indicated by a mobility shift on SDS–PAGE corresponding to the molecular weight of IASD (˜500 Da).
C Protease protection assay in mitochondria expressing Mpc1-GFP, Mpc2-GFP, or Mpc3-GFP. Intact mitochondria (M), mitoplasts with a ruptured outer membrane after hypo-osmotic swelling (Sw), or mitochondrial lysates with 0.5% Triton X-100 (Tr) were treated with proteinase K (PK). Loading controls are Tom70 (outer membrane), Tim23 (inner membrane), and Mdh1 (matrix). GFP-fused Mpc2 and Mpc3 are degraded in mitoplasts, whereas Mpc1-GFP is not.
D Model showing the proposed membrane topology of Mpc1 and Mpc3. Whereas Mpc1 has only two transmembrane helices, Mpc3 has three.
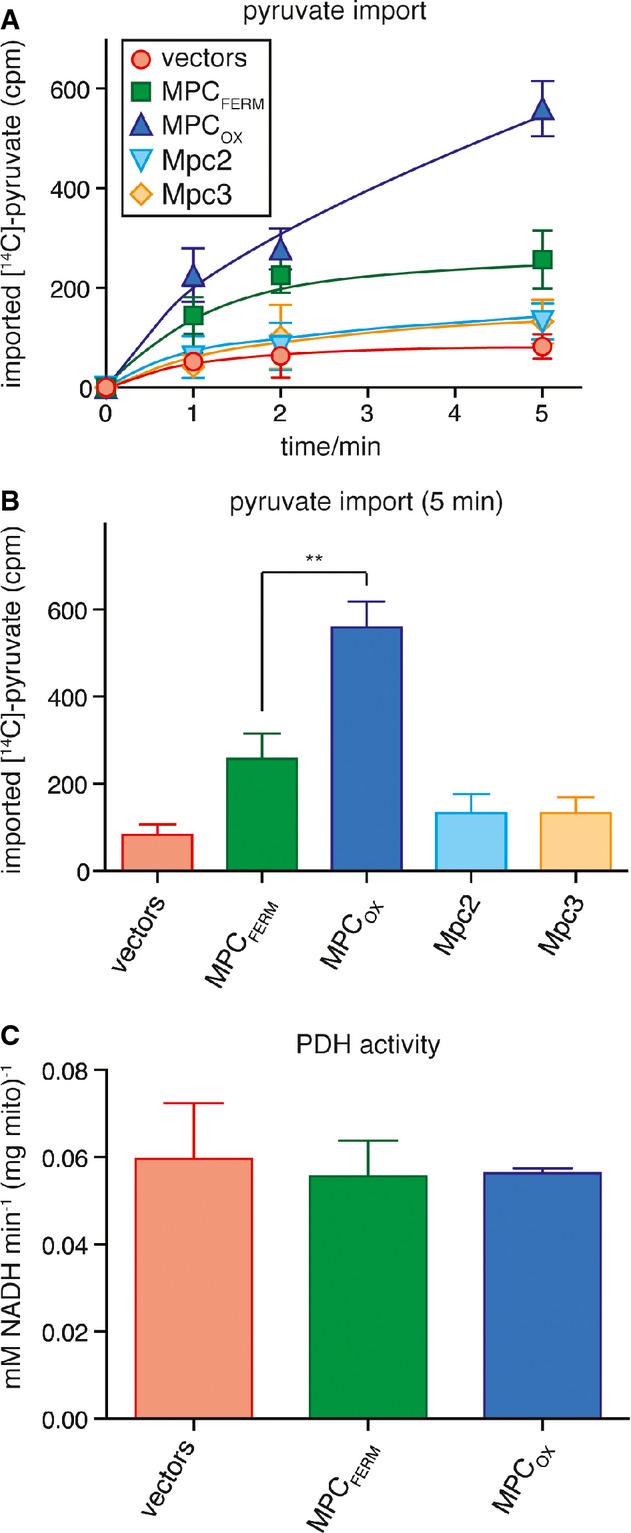
MPCOX has higher transport activity than MPCFERM. The uptake of 14C-labeled pyruvate into intact mitochondria was measured in vitro. Mitochondria had been isolated from cells grown in glycerol-containing medium and expressing no subunit (vectors), MPCFERM, MPCOX, Mpc2, or Mpc3. Imported pyruvate was quantified by re-isolation of mitochondria and subsequent scintillation counting after 1, 2, or 5 min of incubation with [14C]-pyruvate.
Imported [14C]-pyruvate as in (A) after 5-min incubation. The difference between MPCOX and MPCFERM (**P = 0.0037) was significant (unpaired t-test).
Pyruvate dehydrogenase activity was measured in lysates of mitochondria expressing no subunit (vectors), MPCFERM, or MPCOX.
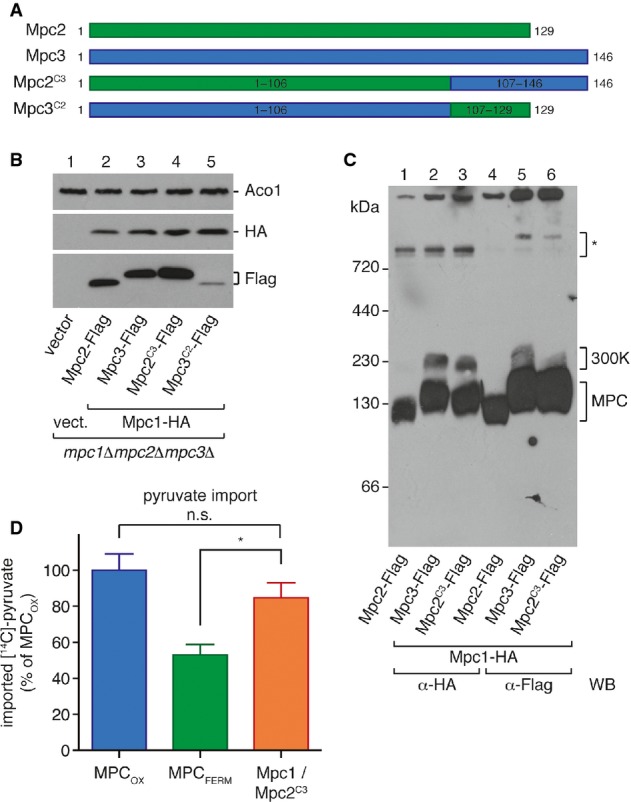
Scheme of the chimeric protein constructs Mpc2C3 and Mpc3C2. The C-terminal region of Mpc2 was replaced by the corresponding region of Mpc3, and vice versa.
The chimeric construct Mpc2C3 is expressed in isolated mitochondria, while Mpc3C2 is not. Mpc1-HA and the Flag-tagged chimeric proteins were detected by Western blotting in isolated mitochondria.
Mpc2C3 forms a carrier complex with Mpc1, as analyzed by BN-PAGE. When co-expressed with Mpc1, Mpc2C3 shows the same complex pattern as Mpc3, including the 300K complex.
Pyruvate transport activity of the Mpc1/Mpc2C3 complex reaches ˜85% of MPCOX activity. Uptake of [14C]-pyruvate into mitochondria isolated from yeast cells expressing the indicated MPC protein combinations was measured in vitro. Difference between MPCFERM and Mpc1/Mpc2C3 is significant (unpaired t-test; *P = 0.02). Means of n = 4 experiments are shown, with error bars representing SEM.
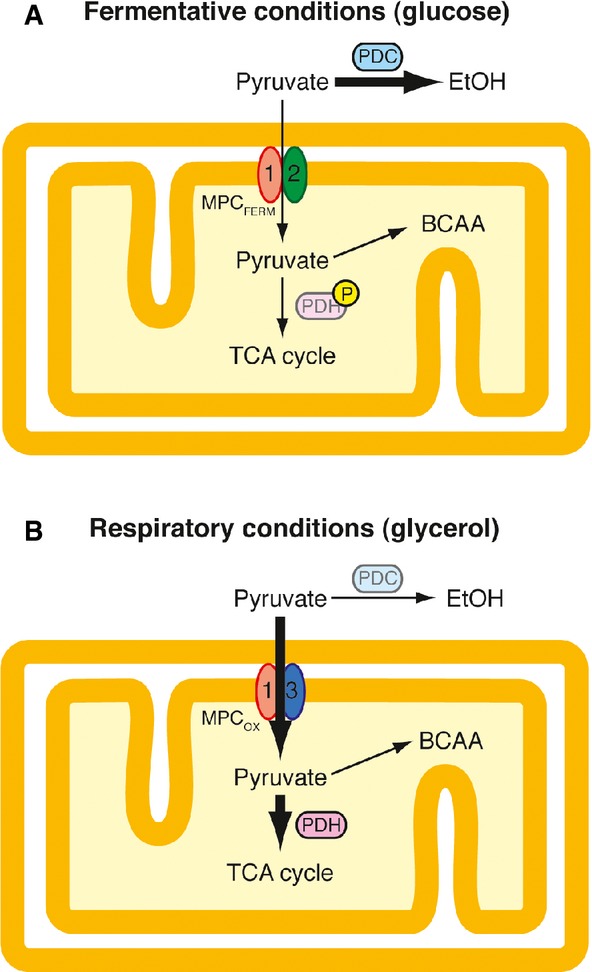
Fermentative conditions. The low-activity MPCFERM (composed of Mpc1 and Mpc2) is competing for cytosolic pyruvate produced through glycolysis with pyruvate decarboxylase (PDC), the first enzyme of the alcoholic fermentation pathway. Consequently, the majority of pyruvate remains in the cytosol is decarboxylated by PDC and is finally transformed into ethanol. Low-level transport of pyruvate into mitochondria is required for the biosynthesis of leucine and valine. In addition, mitochondrial pyruvate oxidation may be blocked by a hypothetical inactivation of pyruvate dehydrogenase (PDH) by phosphorylation of the E1 subunit under these conditions.
Respiratory conditions. The high-activity MPCOX (composed of Mpc1 and Mpc3) is expressed while PDC is downregulated. This allows the efficient uptake of pyruvate into mitochondria. Inside the organelle, pyruvate is converted to acetyl-CoA by PDH, which enters the TCA cycle and thus finally fuels ATP production in the respiratory chain. The subunit exchange in the MPC thus very likely participates in the Crabtree effect in Saccharomyces cerevisiae.
Comment in
-
Metabolic remodeling: a pyruvate transport affair.EMBO J. 2015 Apr 1;34(7):835-7. doi: 10.15252/embj.201591228. Epub 2015 Feb 27. EMBO J. 2015. PMID: 25725020 Free PMC article.
References
-
- de Assis LJ, Zingali RB, Masuda CA, Rodrigues SP, Montero-Lomeli M. Pyruvate decarboxylase activity is regulated by the Ser/Thr protein phosphatase Sit4p in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2013;13:518–528. - PubMed
-
- Bricker DK, Taylor EB, Schell JC, Orsak T, Boutron A, Chen YC, Cox JE, Cardon CM, Van Vranken JG, Dephoure N, Redin C, Boudina S, Gygi SP, Brivet M, Thummel CS, Rutter J. A mitochondrial pyruvate carrier required for pyruvate uptake in yeast, Drosophila, and humans. Science. 2012;337:96–100. - PMC - PubMed
-
- De Deken RH. The Crabtree effect: a regulatory system in yeast. J Gen Microbiol. 1966;44:149–156. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
