

The glycosylation status of PrPC is a key factor in determining transmissible spongiform encephalopathy transmission between species
- PMID: 25673720
- PMCID: PMC4403468
- DOI: 10.1128/JVI.02296-14
The glycosylation status of PrPC is a key factor in determining transmissible spongiform encephalopathy transmission between species
Abstract
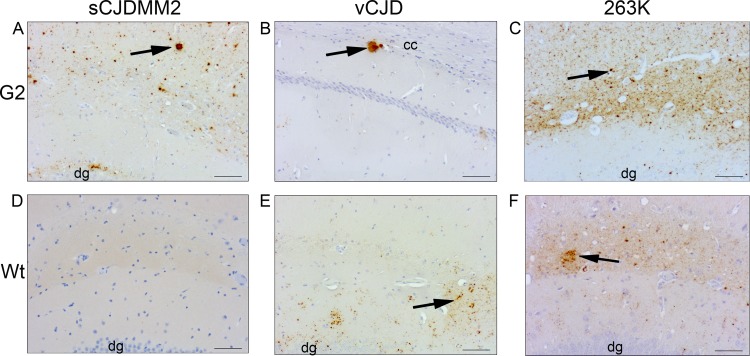
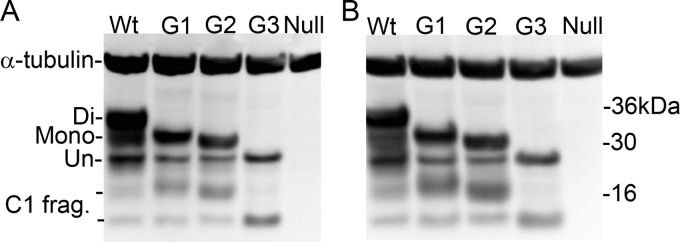
The risk of transmission of transmissible spongiform encephalopathies (TSE) between different species has been notoriously unpredictable because the mechanisms of transmission are not fully understood. A transmission barrier between species often prevents infection of a new host with a TSE agent. Nonetheless, some TSE agents are able to cross this barrier and infect new species, with devastating consequences. The host PrP(C) misfolds during disease pathogenesis and has a major role in controlling the transmission of agents between species, but sequence compatibility between host and agent PrP(C) does not fully explain host susceptibility. PrP(C) is posttranslationally modified by the addition of glycan moieties which have an important role in the infectious process. Here, we show in vivo that glycosylation of the host PrP(C) has a significant impact on the transmission of TSE between different host species. We infected mice carrying different glycosylated forms of PrP(C) with two human agents (sCJDMM2 and vCJD) and one hamster strain (263K). The absence of glycosylation at both or the first PrP(C) glycosylation site in the host results in almost complete resistance to disease. The absence of the second site of N-glycan has a dramatic effect on the barrier to transmission between host species, facilitating the transmission of sCJDMM2 to a host normally resistant to this agent. These results highlight glycosylation of PrP(C) as a key factor in determining the transmission efficiency of TSEs between different species.
Importance: The risks of transmission of TSE between different species are difficult to predict due to a lack of knowledge over the mechanisms of disease transmission; some strains of TSE are able to cross a species barrier, while others do not. The host protein, PrP(C), plays a major role in disease transmission. PrP(C) undergoes posttranslational glycosylation, and the addition of these glycans may play a role in disease transmission. We infected mice that express different forms of glycosylated PrP(C) with three different TSE agents. We demonstrate that changing the glycosylation status of the host can have profound effects on disease transmission, changing host susceptibility and incubation times. Our results show that PrP(C) glycosylation is a key factor in determining risks of TSE transmission between species.
Copyright © 2015, Wiseman et al.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- 069283/WT_/Wellcome Trust/United Kingdom
- BBS/E/D/20251967/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/D/20251968/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- MR/L016400/1/MRC_/Medical Research Council/United Kingdom
- G0900580/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
