

Anti-V3/Glycan and Anti-MPER Neutralizing Antibodies, but Not Anti-V2/Glycan Site Antibodies, Are Strongly Associated with Greater Anti-HIV-1 Neutralization Breadth and Potency
- PMID: 25673728
- PMCID: PMC4442498
- DOI: 10.1128/JVI.00129-15
Anti-V3/Glycan and Anti-MPER Neutralizing Antibodies, but Not Anti-V2/Glycan Site Antibodies, Are Strongly Associated with Greater Anti-HIV-1 Neutralization Breadth and Potency
Abstract
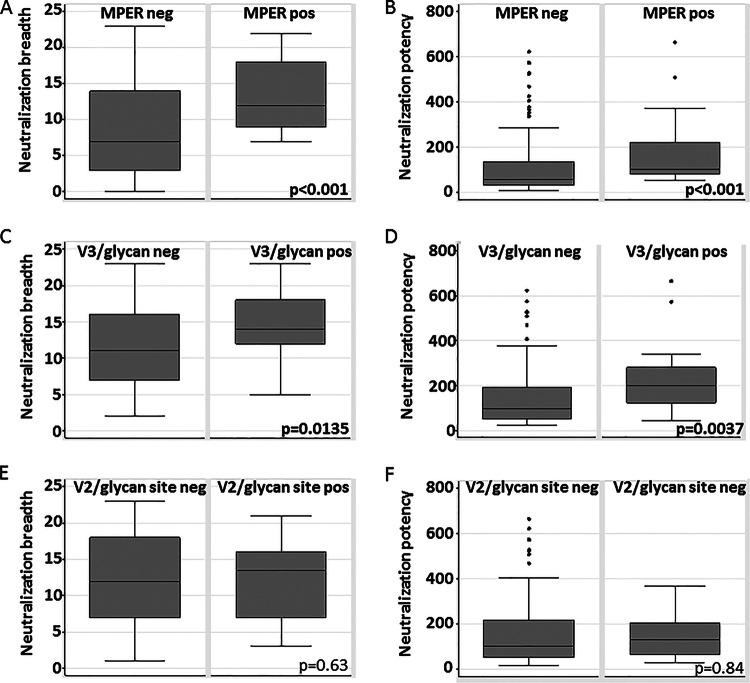
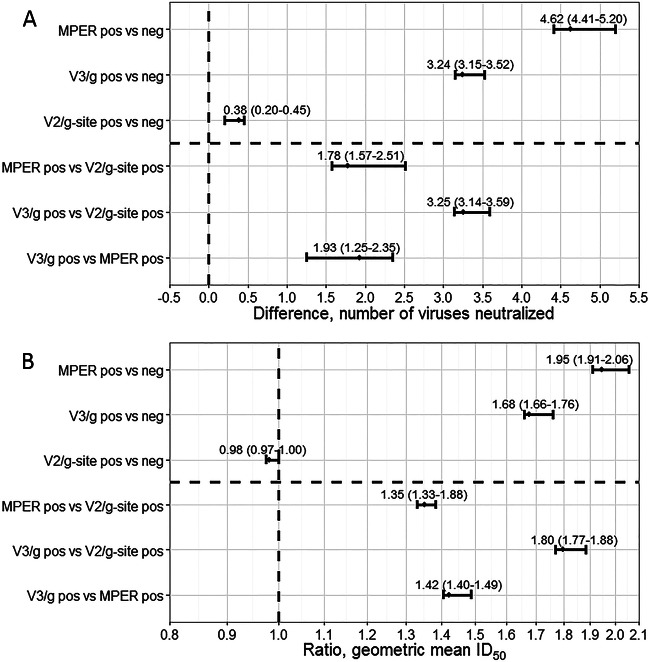
The membrane-proximal external region (MPER), the V2/glycan site (initially defined by PG9 and PG16 antibodies), and the V3/glycans (initially defined by PGT121-128 antibodies) are targets of broadly neutralizing antibodies and potential targets for anti-HIV-1 antibody-based vaccines. Recent evidence shows that antibodies with moderate neutralization breadth are frequently attainable, with 50% of sera from chronically infected individuals neutralizing ≥ 50% of a large, diverse set of viruses. Nonetheless, there is little systematic information addressing which specificities are preferentially targeted among such commonly found, moderately broadly neutralizing sera. We explored associations between neutralization breadth and potency and the presence of neutralizing antibodies targeting the MPER, V2/glycan site, and V3/glycans in sera from 177 antiretroviral-naive HIV-1-infected (>1 year) individuals. Recognition of both MPER and V3/glycans was associated with increased breadth and potency. MPER-recognizing sera neutralized 4.62 more panel viruses than MPER-negative sera (95% prediction interval [95% PI], 4.41 to 5.20), and V3/glycan-recognizing sera neutralized 3.24 more panel viruses than V3/glycan-negative sera (95% PI, 3.15 to 3.52). In contrast, V2/glycan site-recognizing sera neutralized only 0.38 more panel viruses (95% PI, 0.20 to 0.45) than V2/glycan site-negative sera and no association between V2/glycan site recognition and breadth or potency was observed. Despite autoreactivity of many neutralizing antibodies recognizing MPER and V3/glycans, antibodies to these sites are major contributors to neutralization breadth and potency in this cohort. It may therefore be appropriate to focus on developing immunogens based upon the MPER and V3/glycans.
Importance: Previous candidate HIV vaccines have failed either to induce wide-coverage neutralizing antibodies or to substantially protect vaccinees. Therefore, current efforts focus on novel approaches never before successfully used in vaccine design, including modeling epitopes. Candidate immunogen models identified by broadly neutralizing antibodies include the membrane-proximal external region (MPER), V3/glycans, and the V2/glycan site. Autoreactivity and polyreactivity of anti-MPER and anti-V3/glycan antibodies are thought to pose both direct and indirect barriers to achieving neutralization breadth. We found that antibodies to the MPER and the V3/glycans contribute substantially to neutralization breadth and potency. In contrast, antibodies to the V2/glycan site were not associated with neutralization breadth/potency. This suggests that the autoreactivity effect is not critical and that the MPER and the V3/glycans should remain high-priority vaccine candidates. The V2/glycan site result is surprising because broadly neutralizing antibodies to this site have been repeatedly observed. Vaccine design priorities should shift toward the MPER and V3/glycans.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Burton DR, Ahmed R, Barouch DH, Butera ST, Crotty S, Godzik A, Kaufmann DE, McElrath MJ, Nussenzweig MC, Pulendran B, Scanlan CN, Schief WR, Silvestri G, Streeck H, Walker BD, Walker LM, Ward AB, Wilson IA, Wyatt R. 2012. A blueprint for HIV vaccine discovery. Cell Host Microbe 12:396–407. doi: 10.1016/j.chom.2012.09.008. - DOI - PMC - PubMed
-
- Huang J, Kang BH, Pancera M, Lee JH, Tong T, Feng Y, Georgiev IS, Chuang GY, Druz A, Doria-Rose NA, Laub L, Sliepen K, van Gils MJ, de la Pena AT, Derking R, Klasse PJ, Migueles SA, Bailer RT, Alam M, Pugach P, Haynes BF, Wyatt RT, Sanders RW, Binley JM, Ward AB, Mascola JR, Kwong PD, Connors M. 2014. Broad and potent HIV-1 neutralization by a human antibody that binds the gp41-gp120 interface. Nature 515:138–142. doi: 10.1038/nature13601. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
