

Functional cooperation between the IP3 receptor and phospholipase C secures the high sensitivity to light of Drosophila photoreceptors in vivo
- PMID: 25673847
- PMCID: PMC4323532
- DOI: 10.1523/JNEUROSCI.3933-14.2015
Functional cooperation between the IP3 receptor and phospholipase C secures the high sensitivity to light of Drosophila photoreceptors in vivo
Abstract
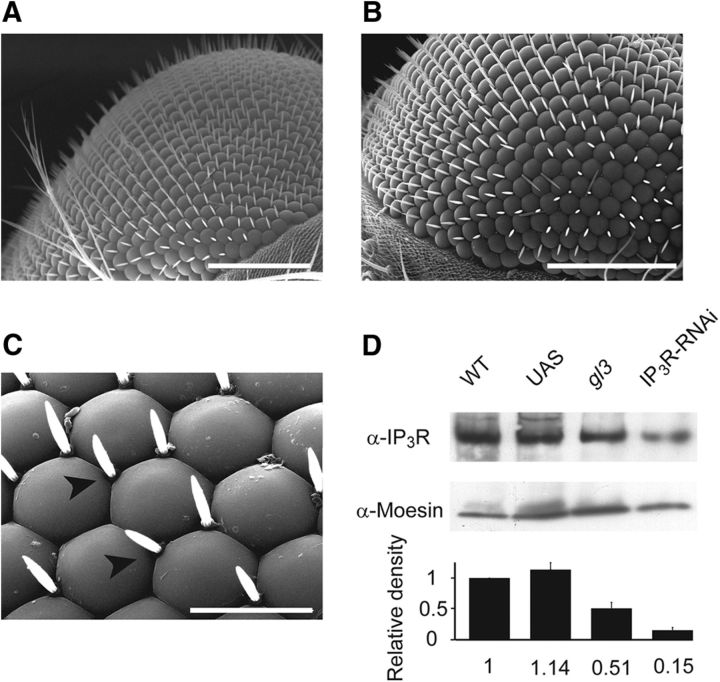
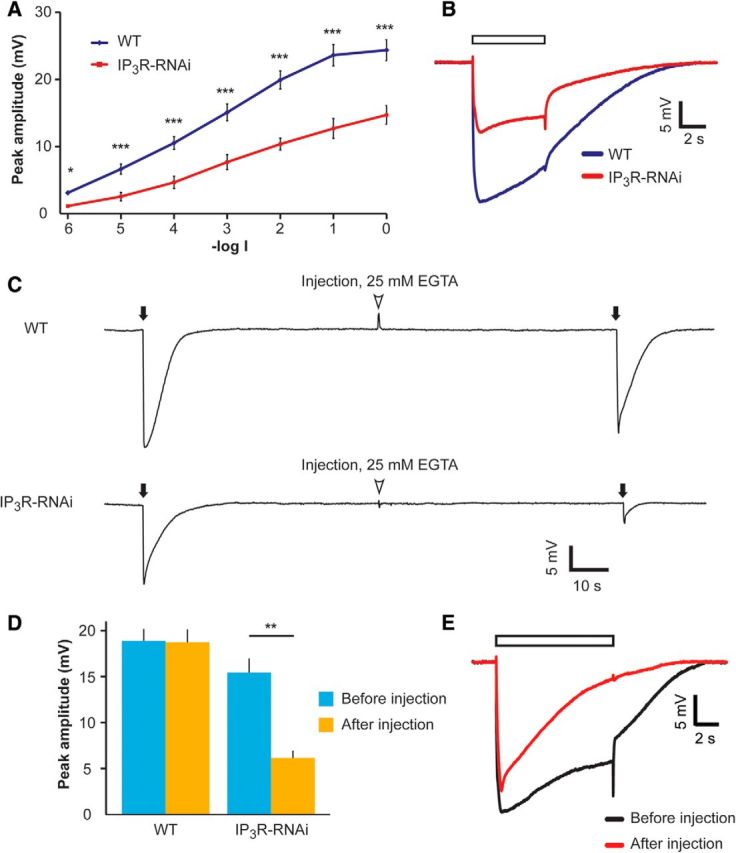
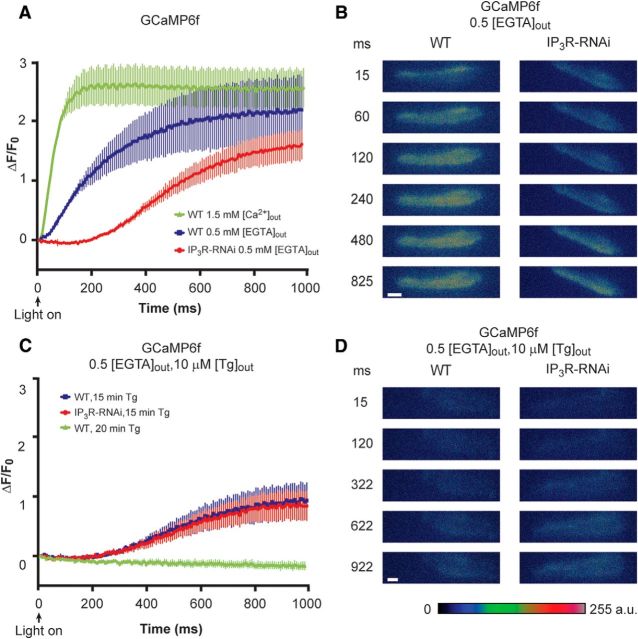
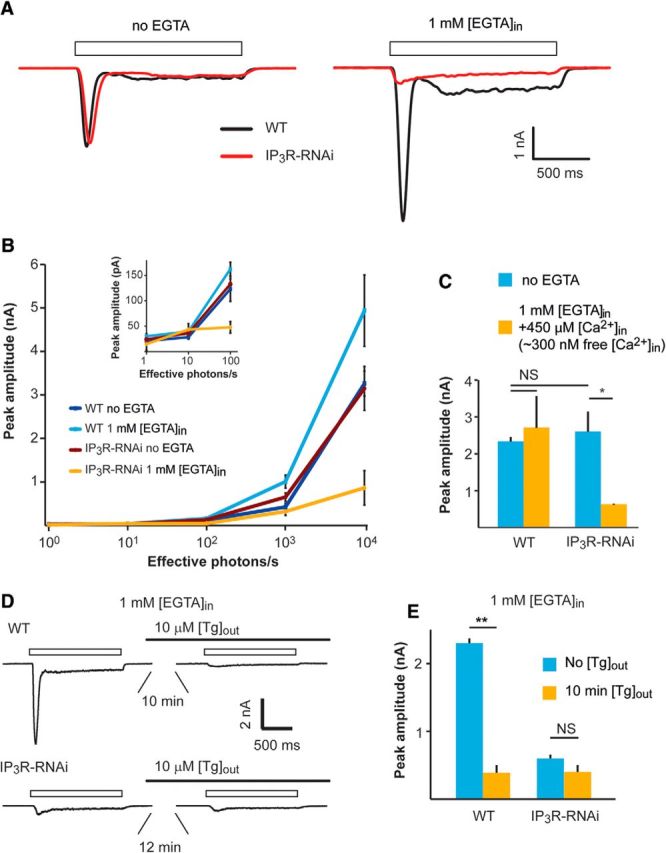
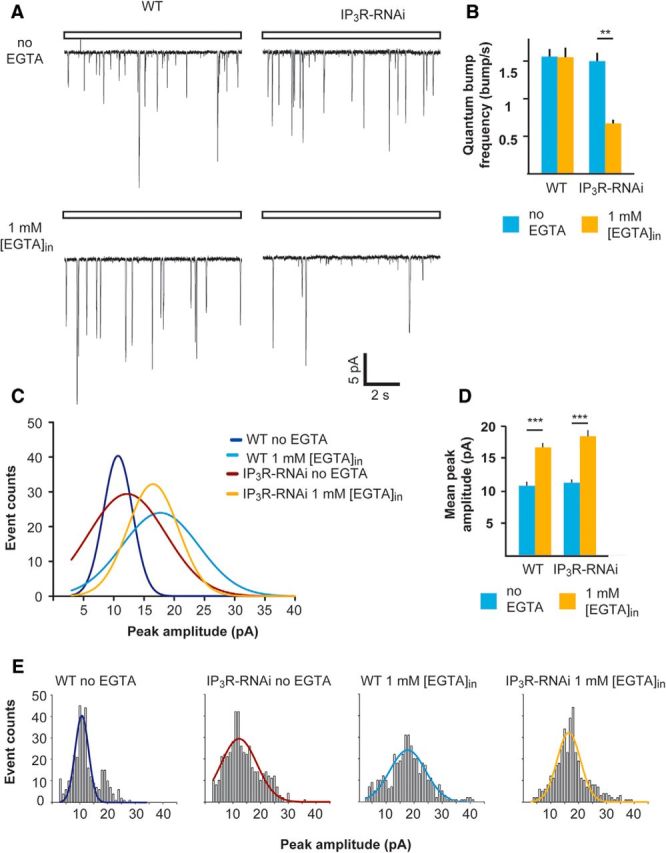
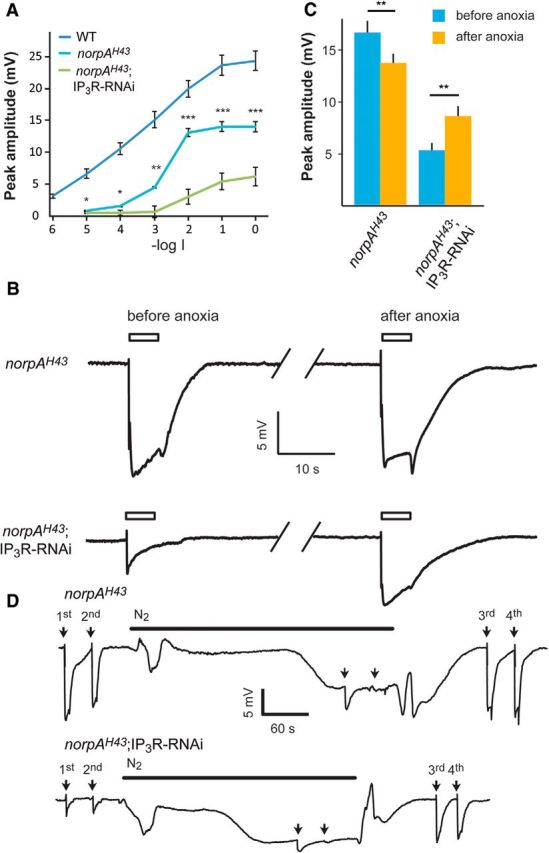
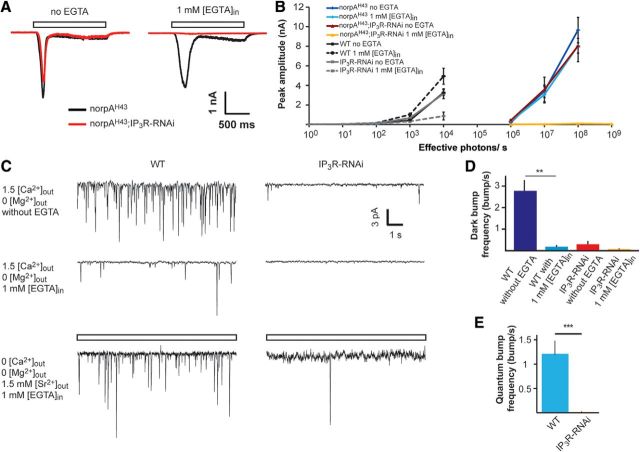
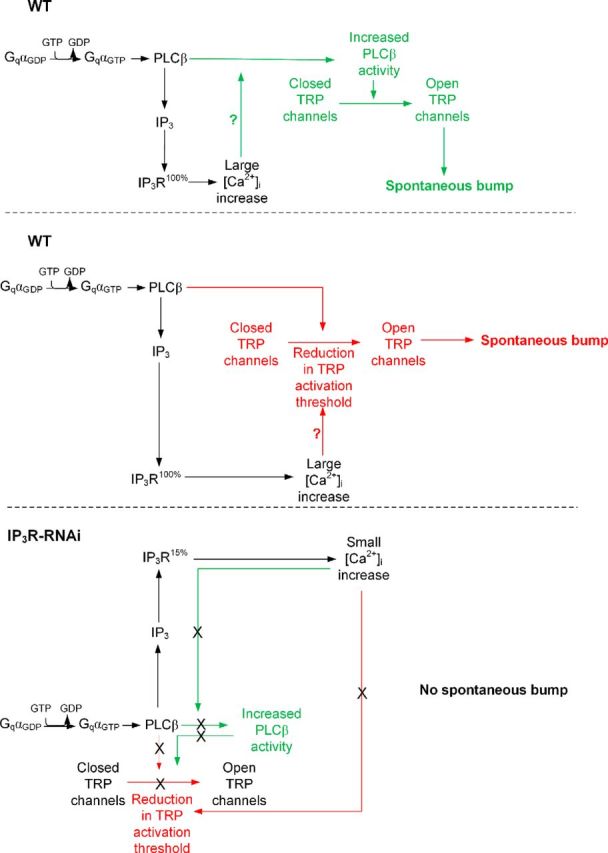
Drosophila phototransduction is a model system for the ubiquitous phosphoinositide signaling. In complete darkness, spontaneous unitary current events (dark bumps) are produced by spontaneous single Gqα activation, while single-photon responses (quantum bumps) arise from synchronous activation of several Gqα molecules. We have recently shown that most of the spontaneous single Gqα activations do not produce dark bumps, because of a critical phospholipase Cβ (PLCβ) activity level required for bump generation. Surpassing the threshold of channel activation depends on both PLCβ activity and cellular [Ca(2+)], which participates in light excitation via a still unclear mechanism. We show here that in IP3 receptor (IP3R)-deficient photoreceptors, both light-activated Ca(2+) release from internal stores and light sensitivity were strongly attenuated. This was further verified by Ca(2+) store depletion, linking Ca(2+) release to light excitation. In IP3R-deficient photoreceptors, dark bumps were virtually absent and the quantum-bump rate was reduced, indicating that Ca(2+) release from internal stores is necessary to reach the critical level of PLCβ catalytic activity and the cellular [Ca(2+)] required for excitation. Combination of IP3R knockdown with reduced PLCβ catalytic activity resulted in highly suppressed light responses that were partially rescued by cellular Ca(2+) elevation, showing a functional cooperation between IP3R and PLCβ via released Ca(2+). These findings suggest that in contrast to the current dogma that Ca(2+) release via IP3R does not participate in light excitation, we show that released Ca(2+) plays a critical role in light excitation. The positive feedback between PLCβ and IP3R found here may represent a common feature of the inositol-lipid signaling.
Keywords: Ca2+ release; Drosophila; IP3 receptor; phospholipase C; photoreceptors; phototransduction.
Copyright © 2015 the authors 0270-6474/15/352530-17$15.00/0.
Figures
Similar articles
-
Phototransduction in Drosophila Is Compromised by Gal4 Expression but not by InsP3 Receptor Knockdown or Mutation.eNeuro. 2017 Jun 26;4(3):ENEURO.0143-17.2017. doi: 10.1523/ENEURO.0143-17.2017. eCollection 2017 May-Jun. eNeuro. 2017. PMID: 28660247 Free PMC article.
-
Phospholipase C-mediated suppression of dark noise enables single-photon detection in Drosophila photoreceptors.J Neurosci. 2012 Feb 22;32(8):2722-33. doi: 10.1523/JNEUROSCI.5221-11.2012. J Neurosci. 2012. PMID: 22357856 Free PMC article.
-
A common mechanism underlies vertebrate calcium signaling and Drosophila phototransduction.J Neurosci. 2001 Apr 15;21(8):2622-9. doi: 10.1523/JNEUROSCI.21-08-02622.2001. J Neurosci. 2001. PMID: 11306615 Free PMC article.
-
The Drosophila light-activated TRP and TRPL channels - Targets of the phosphoinositide signaling cascade.Prog Retin Eye Res. 2018 Sep;66:200-219. doi: 10.1016/j.preteyeres.2018.05.001. Epub 2018 May 5. Prog Retin Eye Res. 2018. PMID: 29738822 Review.
-
Phosphoinositide-mediated phototransduction in Drosophila photoreceptors: the role of Ca2+ and trp.Cell Calcium. 1995 Oct;18(4):256-74. doi: 10.1016/0143-4160(95)90023-3. Cell Calcium. 1995. PMID: 8556766 Review.
Cited by
-
A Comparative Perspective on Functionally-Related, Intracellular Calcium Channels: The Insect Ryanodine and Inositol 1,4,5-Trisphosphate Receptors.Biomolecules. 2021 Jul 15;11(7):1031. doi: 10.3390/biom11071031. Biomolecules. 2021. PMID: 34356655 Free PMC article. Review.
-
Application of Fluorescent Proteins for Functional Dissection of the Drosophila Visual System.Int J Mol Sci. 2021 Aug 19;22(16):8930. doi: 10.3390/ijms22168930. Int J Mol Sci. 2021. PMID: 34445636 Free PMC article. Review.
-
Phototransduction in Drosophila Is Compromised by Gal4 Expression but not by InsP3 Receptor Knockdown or Mutation.eNeuro. 2017 Jun 26;4(3):ENEURO.0143-17.2017. doi: 10.1523/ENEURO.0143-17.2017. eCollection 2017 May-Jun. eNeuro. 2017. PMID: 28660247 Free PMC article.
-
Rapid Release of Ca2+ from Endoplasmic Reticulum Mediated by Na+/Ca2+ Exchange.J Neurosci. 2020 Apr 15;40(16):3152-3164. doi: 10.1523/JNEUROSCI.2675-19.2020. Epub 2020 Mar 10. J Neurosci. 2020. PMID: 32156830 Free PMC article.
-
The Phosphorylation State of the Drosophila TRP Channel Modulates the Frequency Response to Oscillating Light In Vivo.J Neurosci. 2017 Apr 12;37(15):4213-4224. doi: 10.1523/JNEUROSCI.3670-16.2017. Epub 2017 Mar 17. J Neurosci. 2017. PMID: 28314815 Free PMC article.
References
-
- Abramoff MD, Magelhaes PJ, Ram SJ. Image processing with ImageJ. Biophotonics Int. 2004;11:36–42.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous