

Unraveling the Interaction between FcRn and Albumin: Opportunities for Design of Albumin-Based Therapeutics
- PMID: 25674083
- PMCID: PMC4306297
- DOI: 10.3389/fimmu.2014.00682
Unraveling the Interaction between FcRn and Albumin: Opportunities for Design of Albumin-Based Therapeutics
Abstract
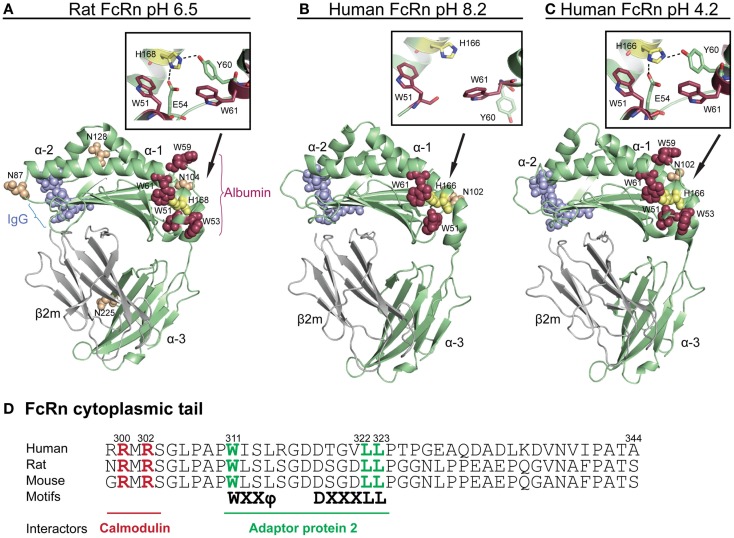
The neonatal Fc receptor (FcRn) was first found to be responsible for transporting antibodies of the immunoglobulin G (IgG) class from the mother to the fetus or neonate as well as for protecting IgG from intracellular catabolism. However, it has now become apparent that the same receptor also binds albumin and plays a fundamental role in homeostatic regulation of both IgG and albumin, as FcRn is expressed in many different cell types and organs at diverse body sites. Thus, to gain a complete understanding of the biological function of each ligand, and also their distribution in the body, an in-depth characterization of how FcRn binds and regulates the transport of both ligands is necessary. Importantly, such knowledge is also relevant when developing new drugs, as IgG and albumin are increasingly utilized in therapy. This review discusses our current structural and biological understanding of the relationship between FcRn and its ligands, with a particular focus on albumin and design of albumin-based therapeutics.
Keywords: FcRn; IgG; albumin; albumin-based therapeutics; half-life; recycling; transcytosis.
Figures




References
-
- Waldmann TA, Strober W. Metabolism of immunoglobulins. Prog Allergy (1969) 13:1–110. - PubMed
-
- Peters T. All about albumin: Biochemistry, Genetics and Medical Applications. San Diego CA: Academic Press; (1996).
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources