

Transcriptome and metabolome analysis of plant sulfate starvation and resupply provides novel information on transcriptional regulation of metabolism associated with sulfur, nitrogen and phosphorus nutritional responses in Arabidopsis
- PMID: 25674096
- PMCID: PMC4309162
- DOI: 10.3389/fpls.2014.00805
Transcriptome and metabolome analysis of plant sulfate starvation and resupply provides novel information on transcriptional regulation of metabolism associated with sulfur, nitrogen and phosphorus nutritional responses in Arabidopsis
Abstract
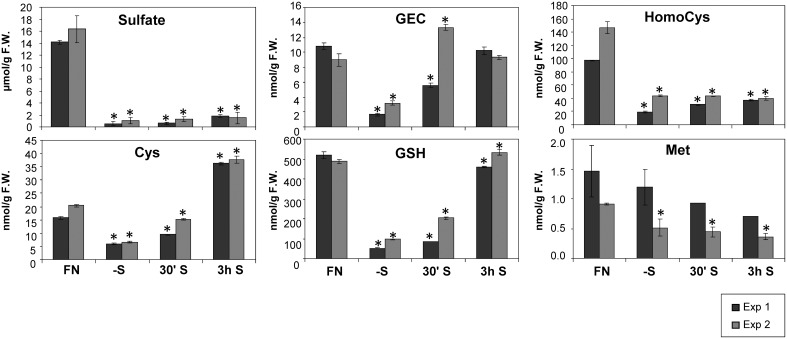
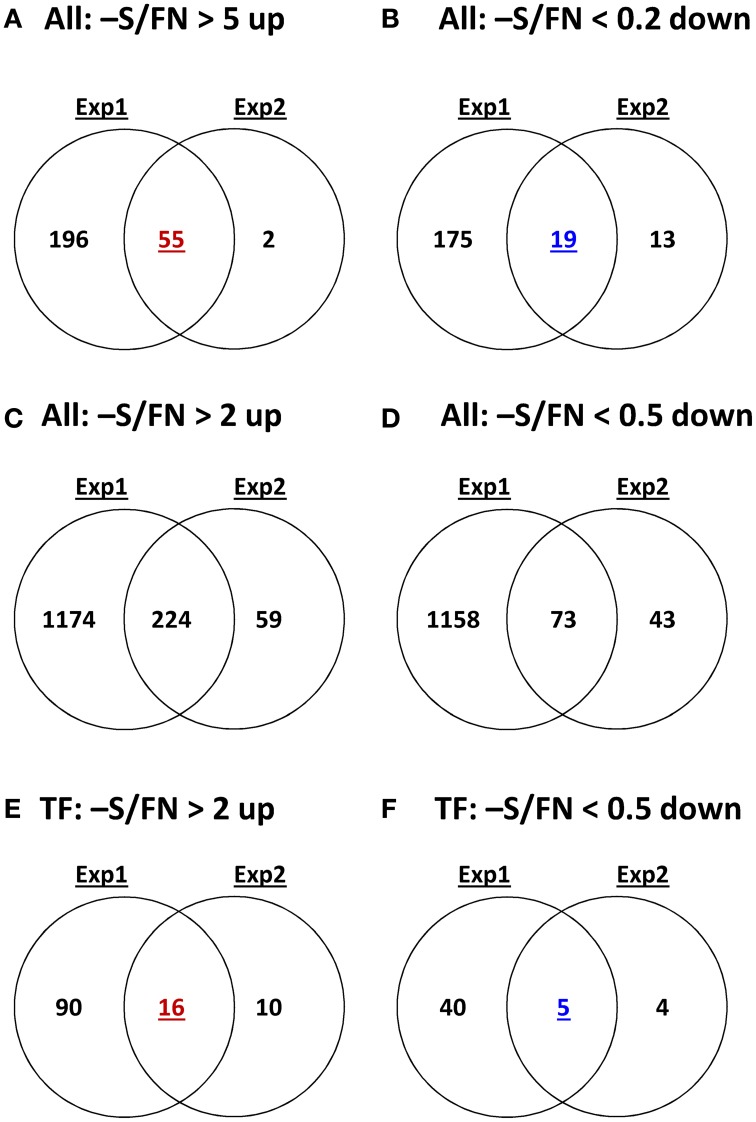
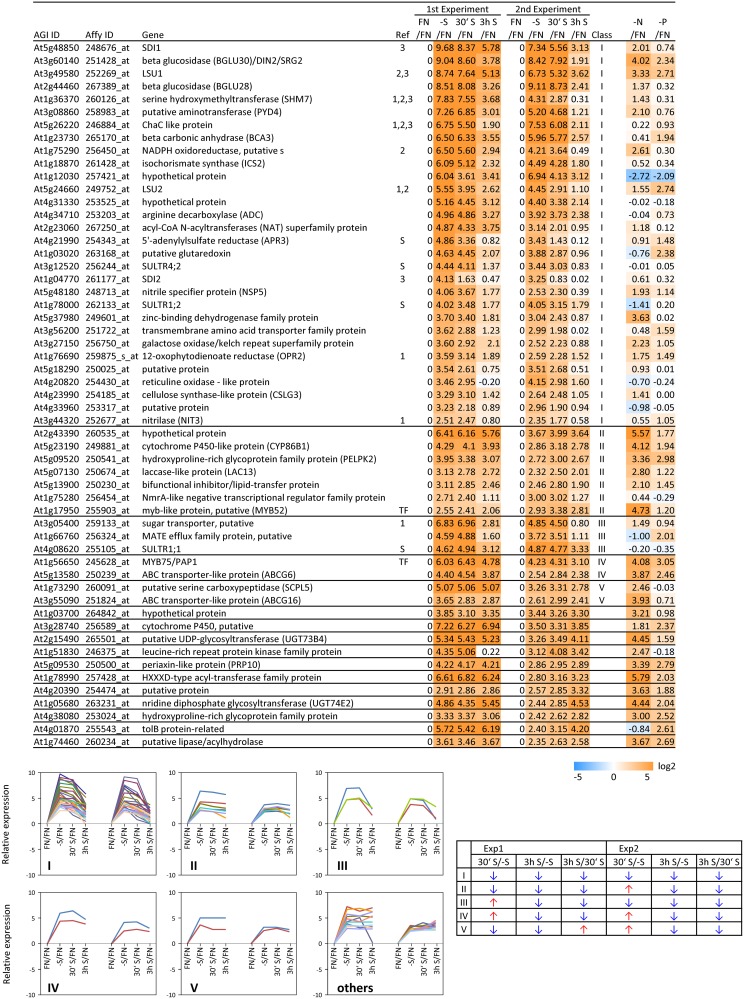
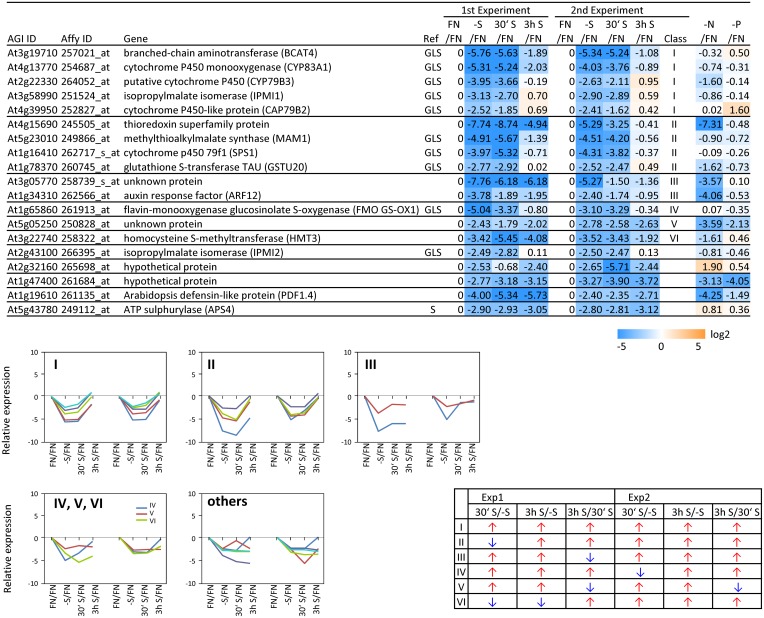
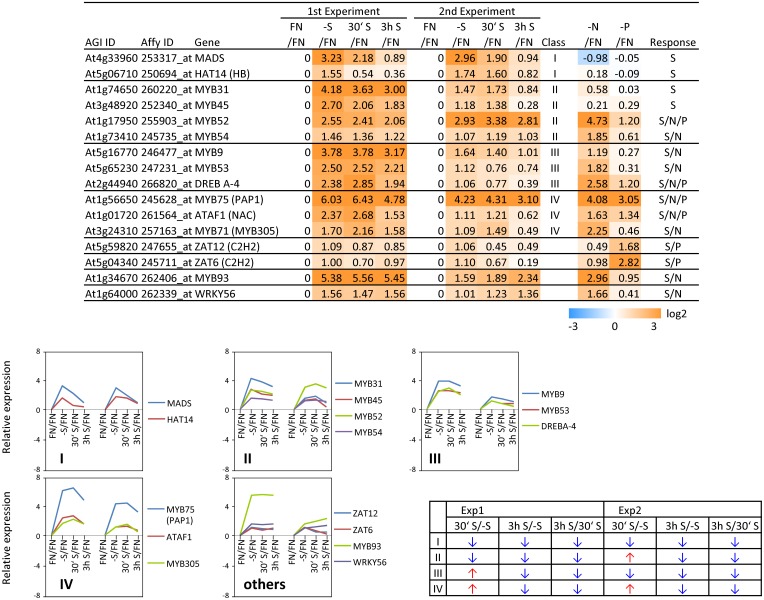
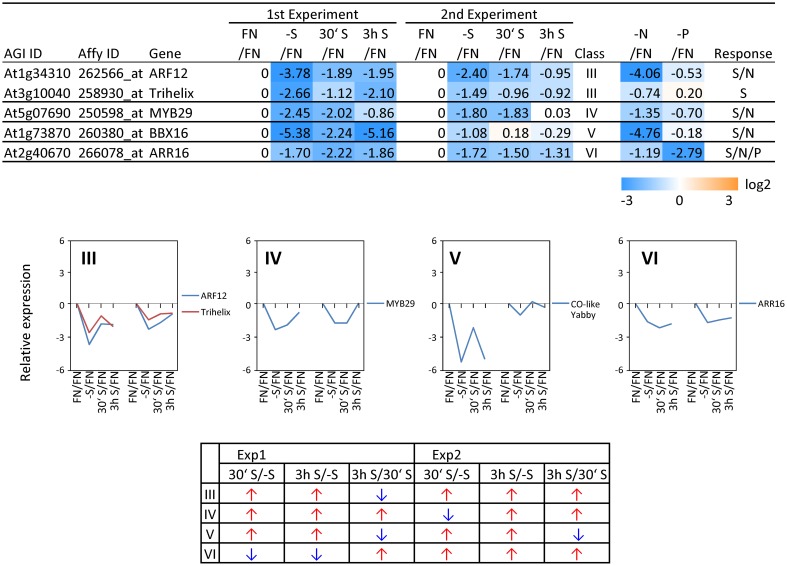
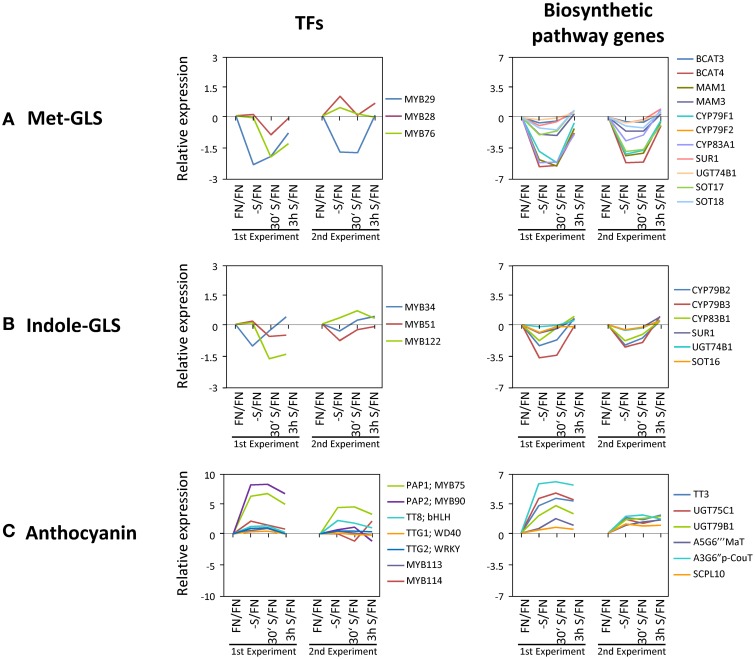
Sulfur is an essential macronutrient for plant growth and development. Reaching a thorough understanding of the molecular basis for changes in plant metabolism depending on the sulfur-nutritional status at the systems level will advance our basic knowledge and help target future crop improvement. Although the transcriptional responses induced by sulfate starvation have been studied in the past, knowledge of the regulation of sulfur metabolism is still fragmentary. This work focuses on the discovery of candidates for regulatory genes such as transcription factors (TFs) using 'omics technologies. For this purpose a short term sulfate-starvation/re-supply approach was used. ATH1 microarray studies and metabolite determinations yielded 21 TFs which responded more than 2-fold at the transcriptional level to sulfate starvation. Categorization by response behaviors under sulfate-starvation/re-supply and other nutrient starvations such as nitrate and phosphate allowed determination of whether the TF genes are specific for or common between distinct mineral nutrient depletions. Extending this co-behavior analysis to the whole transcriptome data set enabled prediction of putative downstream genes. Additionally, combinations of transcriptome and metabolome data allowed identification of relationships between TFs and downstream responses, namely, expression changes in biosynthetic genes and subsequent metabolic responses. Effect chains on glucosinolate and polyamine biosynthesis are discussed in detail. The knowledge gained from this study provides a blueprint for an integrated analysis of transcriptomics and metabolomics and application for the identification of uncharacterized genes.
Keywords: metabolomics; microarray; nitrate; phosphate; sulfate starvation; sulfur; transcription factor; transcriptomics.
Figures
References
-
- Blakely L. M., Durham M., Evans T. A., Blakely R. M. (1982). Experimental studies on lateral root formation in radish seedling roots: I. General methods, developmental stages, spontaneous formation of laterals. Bot. Gaz. 143, 341–352. 10.1086/337308 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous