

In-cell SHAPE reveals that free 30S ribosome subunits are in the inactive state
- PMID: 25675474
- PMCID: PMC4345610
- DOI: 10.1073/pnas.1411514112
In-cell SHAPE reveals that free 30S ribosome subunits are in the inactive state
Abstract
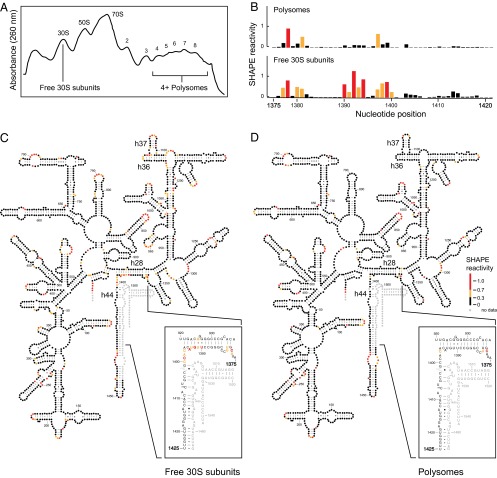
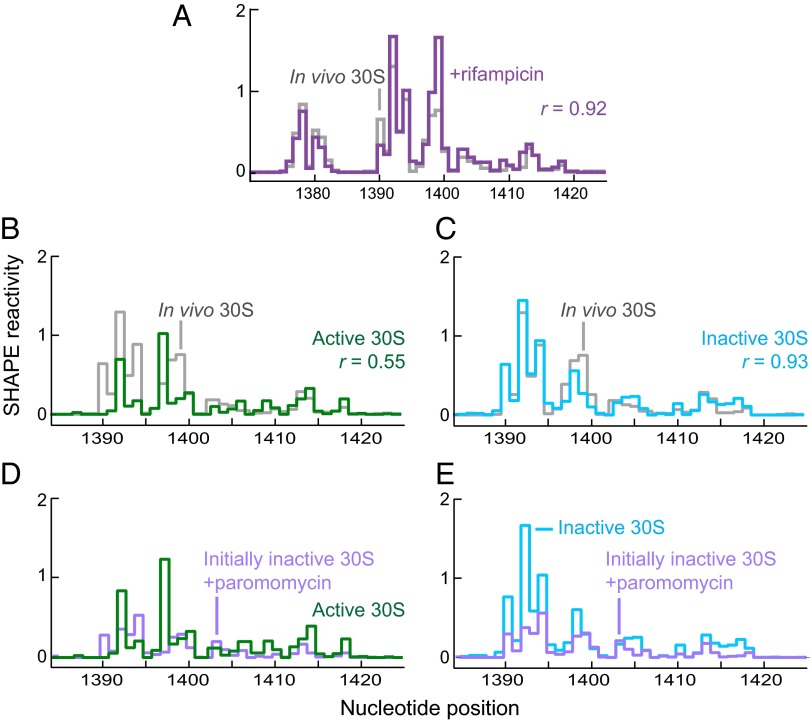
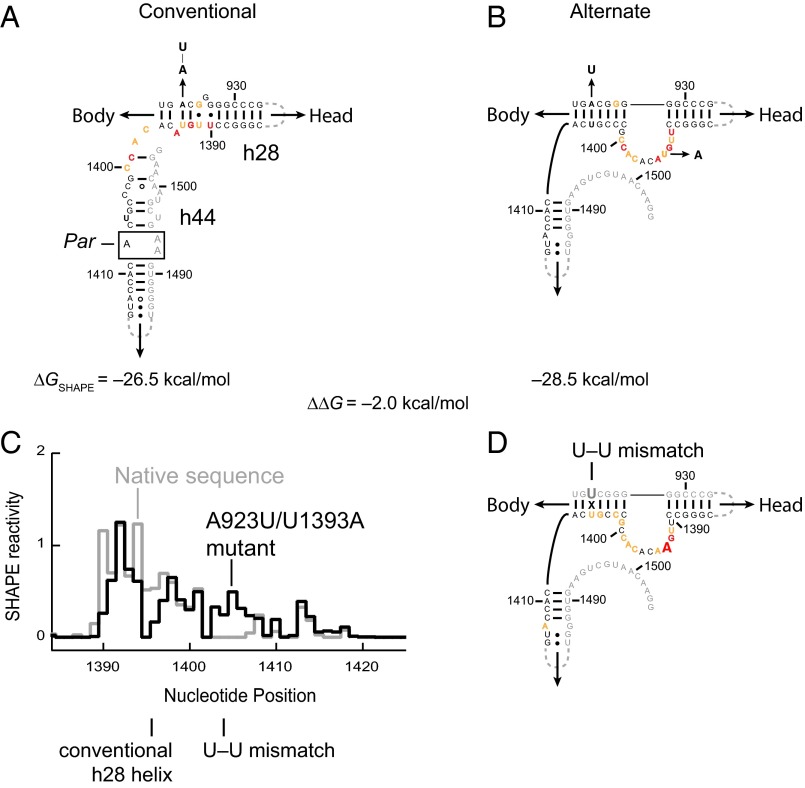
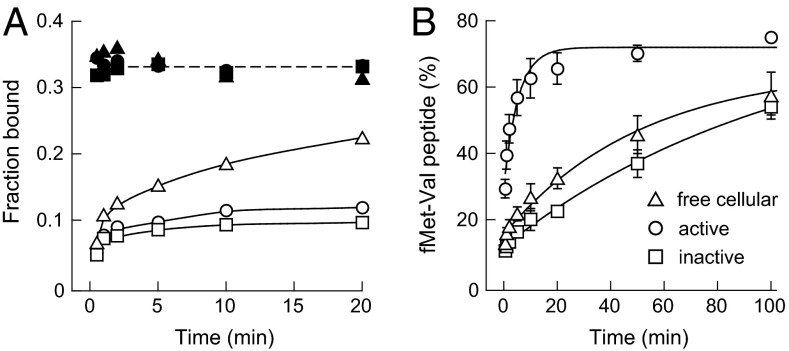
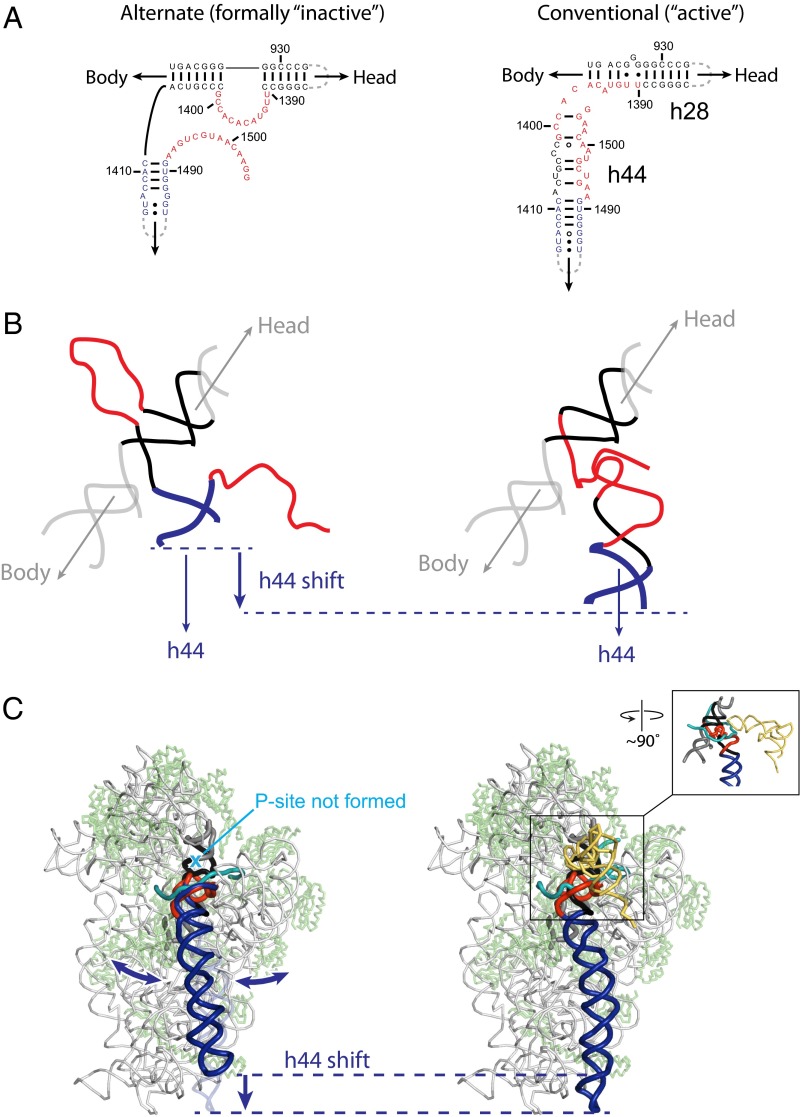
It was shown decades ago that purified 30S ribosome subunits readily interconvert between "active" and "inactive" conformations in a switch that involves changes in the functionally important neck and decoding regions. However, the physiological significance of this conformational change had remained unknown. In exponentially growing Escherichia coli cells, RNA SHAPE probing revealed that 16S rRNA largely adopts the inactive conformation in stably assembled, mature 30S subunits and the active conformation in translating (70S) ribosomes. Inactive 30S subunits bind mRNA as efficiently as active subunits but initiate translation more slowly. Mutations that inhibited interconversion between states compromised translation in vivo. Binding by the small antibiotic paromomycin induced the inactive-to-active conversion, consistent with a low-energy barrier between the two states. Despite the small energetic barrier between states, but consistent with slow translation initiation and a functional role in vivo, interconversion involved large-scale changes in structure in the neck region that likely propagate across the 30S body via helix 44. These findings suggest the inactive state is a biologically relevant alternate conformation that regulates ribosome function as a conformational switch.
Keywords: 16S rRNA; SHAPE; conformational change; in vivo; ribosome.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Zamir A, Miskin R, Elson D. Interconversions between inactive and active forms of ribosomal subunits. FEBS Lett. 1969;3(1):85–88. - PubMed
-
- Moazed D, Van Stolk BJ, Douthwaite S, Noller HF. Interconversion of active and inactive 30 S ribosomal subunits is accompanied by a conformational change in the decoding region of 16 S rRNA. J Mol Biol. 1986;191(3):483–493. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
