

Thermodynamic mechanism for inhibition of lactose permease by the phosphotransferase protein IIAGlc
- PMID: 25675534
- PMCID: PMC4345578
- DOI: 10.1073/pnas.1500891112
Thermodynamic mechanism for inhibition of lactose permease by the phosphotransferase protein IIAGlc
Abstract
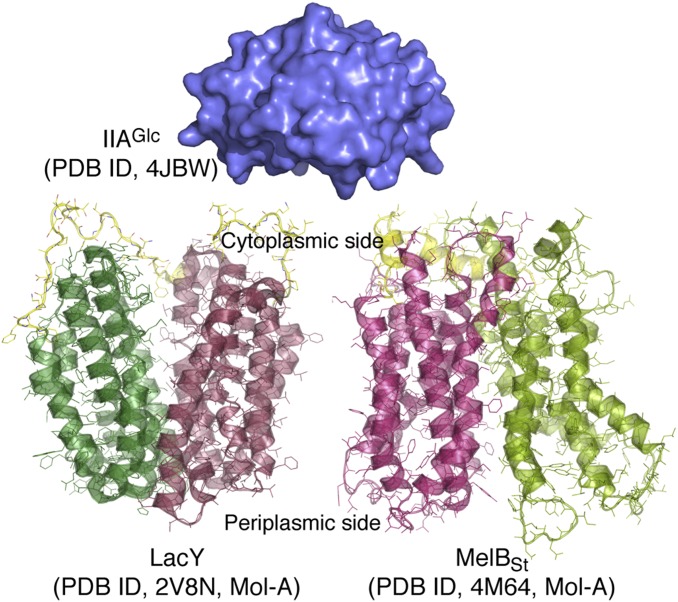
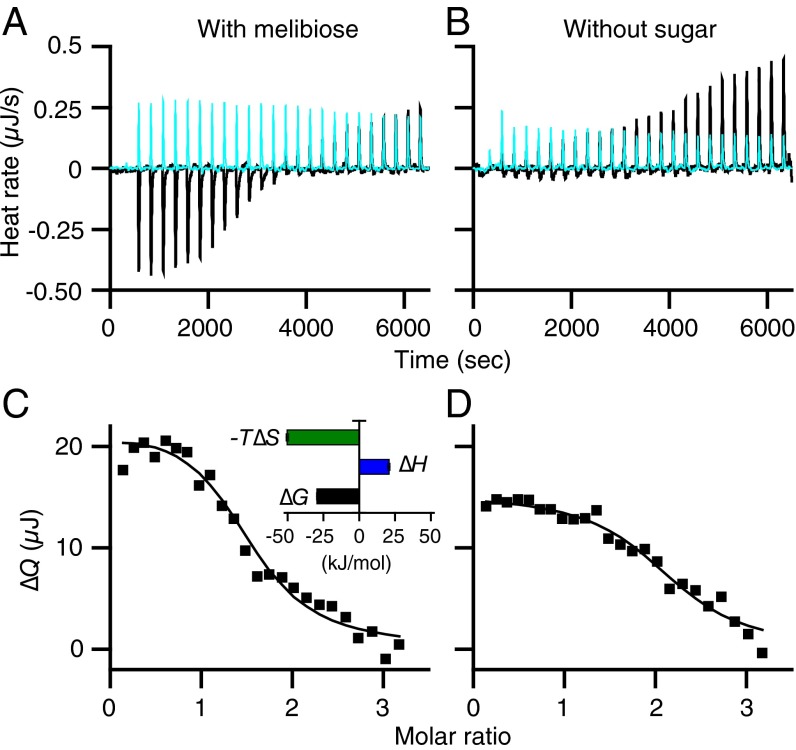
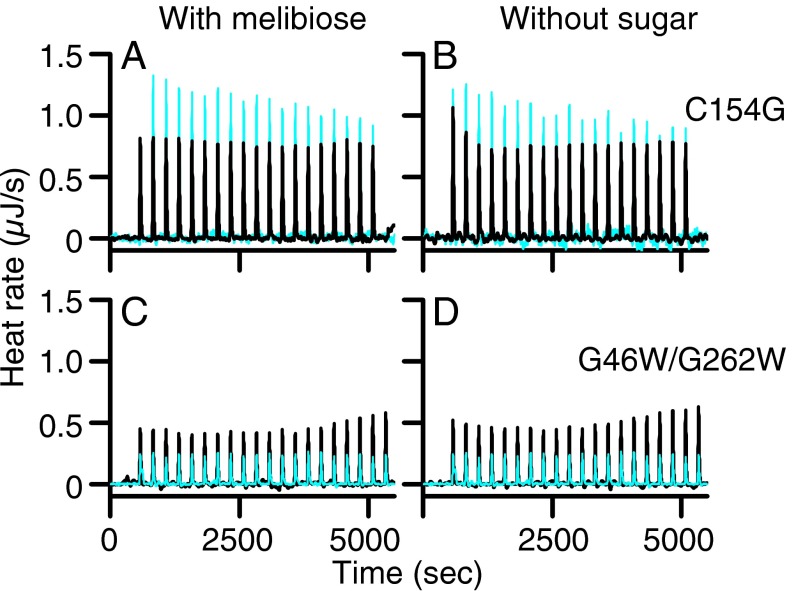
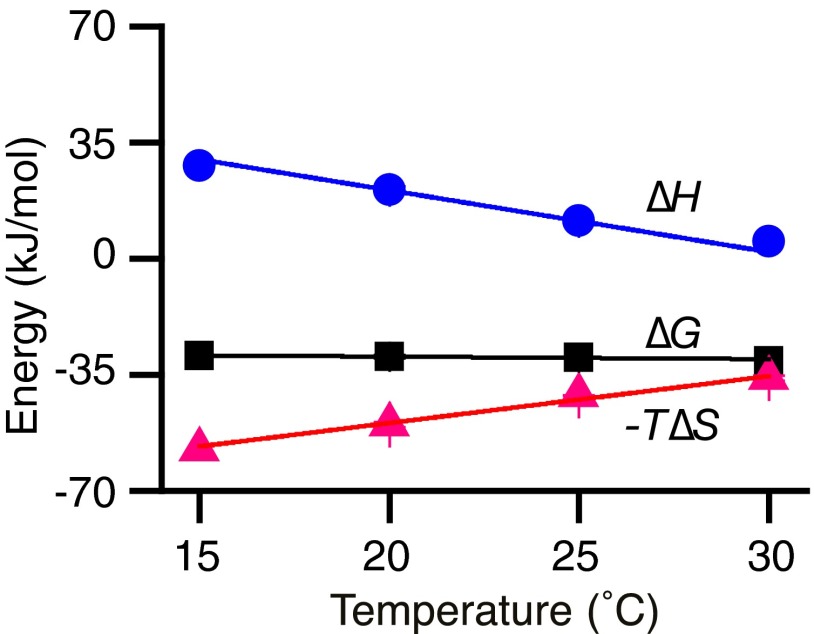
In a variety of bacteria, the phosphotransferase protein IIA(Glc) plays a key regulatory role in catabolite repression in addition to its role in the vectorial phosphorylation of glucose catalyzed by the phosphoenolpyruvate:carbohydrate phosphotransferase system (PTS). The lactose permease (LacY) of Escherichia coli catalyzes stoichiometric symport of a galactoside with an H(+), using a mechanism in which sugar- and H(+)-binding sites become alternatively accessible to either side of the membrane. Both the expression (via regulation of cAMP levels) and the activity of LacY are subject to regulation by IIA(Glc) (inducer exclusion). Here we report the thermodynamic features of the IIA(Glc)-LacY interaction as measured by isothermal titration calorimetry (ITC). The studies show that IIA(Glc) binds to LacY with a Kd of about 5 μM and a stoichiometry of unity and that binding is driven by solvation entropy and opposed by enthalpy. Upon IIA(Glc) binding, the conformational entropy of LacY is restrained, which leads to a significant decrease in sugar affinity. By suppressing conformational dynamics, IIA(Glc) blocks inducer entry into cells and favors constitutive glucose uptake and utilization. Furthermore, the studies support the notion that sugar binding involves an induced-fit mechanism that is inhibited by IIA(Glc) binding. The precise mechanism of the inhibition of LacY by IIA(Glc) elucidated by ITC differs from the inhibition of melibiose permease (MelB), supporting the idea that permeases can differ in their thermodynamic response to binding IIA(Glc).
Keywords: ITC; PTS; protein conformation; protein–protein interactions; sugar/cation symport.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Insights into the inhibitory mechanisms of the regulatory protein IIA(Glc) on melibiose permease activity.J Biol Chem. 2014 Nov 21;289(47):33012-9. doi: 10.1074/jbc.M114.609255. Epub 2014 Oct 8. J Biol Chem. 2014. PMID: 25296751 Free PMC article.
-
Binding of enzyme IIAGlc, a component of the phosphoenolpyruvate:sugar phosphotransferase system, to the Escherichia coli lactose permease.Biochemistry. 2002 Apr 30;41(17):5556-65. doi: 10.1021/bi011990j. Biochemistry. 2002. PMID: 11969416
-
Cooperative binding of the sugar substrates and allosteric regulatory protein (enzyme IIIGlc of the phosphotransferase system) to the lactose and melibiose permeases in Escherichia coli and Salmonella typhimurium.J Bacteriol. 1983 Sep;155(3):1351-7. doi: 10.1128/jb.155.3.1351-1357.1983. J Bacteriol. 1983. PMID: 6350268 Free PMC article.
-
Regulation of galactoside transport by the PTS.J Mol Microbiol Biotechnol. 2001 Jul;3(3):381-4. J Mol Microbiol Biotechnol. 2001. PMID: 11361068 Review.
-
Three-dimensional structures of protein-protein complexes in the E. coli PTS.J Mol Microbiol Biotechnol. 2001 Jul;3(3):347-54. J Mol Microbiol Biotechnol. 2001. PMID: 11361064 Review.
Cited by
-
Flipping the switch: dynamic modulation of membrane transporter activity in bacteria.Microbiology (Reading). 2023 Nov;169(11):001412. doi: 10.1099/mic.0.001412. Microbiology (Reading). 2023. PMID: 37948297 Free PMC article. Review.
-
Cooperative binding ensures the obligatory melibiose/Na+ cotransport in MelB.J Gen Physiol. 2021 Aug 2;153(8):e202012710. doi: 10.1085/jgp.202012710. Epub 2021 Jun 10. J Gen Physiol. 2021. PMID: 34110360 Free PMC article.
-
Contributions of SpoT Hydrolase, SpoT Synthetase, and RelA Synthetase to Carbon Source Diauxic Growth Transitions in Escherichia coli.Front Microbiol. 2018 Aug 3;9:1802. doi: 10.3389/fmicb.2018.01802. eCollection 2018. Front Microbiol. 2018. PMID: 30123210 Free PMC article.
-
Mobile barrier mechanisms for Na+-coupled symport in an MFS sugar transporter.bioRxiv [Preprint]. 2023 Nov 25:2023.09.18.558283. doi: 10.1101/2023.09.18.558283. bioRxiv. 2023. Update in: Elife. 2024 Feb 21;12:RP92462. doi: 10.7554/eLife.92462. PMID: 37790566 Free PMC article. Updated. Preprint.
-
Variable-Internal-Stores models of microbial growth and metabolism with dynamic allocation of cellular resources.J Math Biol. 2017 Jan;74(1-2):409-445. doi: 10.1007/s00285-016-1030-4. Epub 2016 Jun 6. J Math Biol. 2017. PMID: 27271085 Free PMC article.
References
-
- Kaback HR. The role of the phosphoenolpyruvate-phosphotransferase system in the transport of sugars by isolated membrane preparations of Escherichia coli. J Biol Chem. 1968;243(13):3711–3724. - PubMed
-
- Lengeler JW, Jacobson GR, editors. Postma PW. Am. Soc. Microbiol; Washington, DC: 1996. pp. 1149–1174.
-
- Guan L, Kaback HR. Glucose/sugar transport in bacteria. In: Lennarz WJ, Lane MD, editors. Encyclopedia of Biological Chemistry. 2nd Ed. Elsevier; Oxford: 2013. pp. 387–390.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
