

FOXO1 is regulated by insulin and IGF1 in pituitary gonadotropes
- PMID: 25676570
- PMCID: PMC4363278
- DOI: 10.1016/j.mce.2015.02.006
FOXO1 is regulated by insulin and IGF1 in pituitary gonadotropes
Abstract
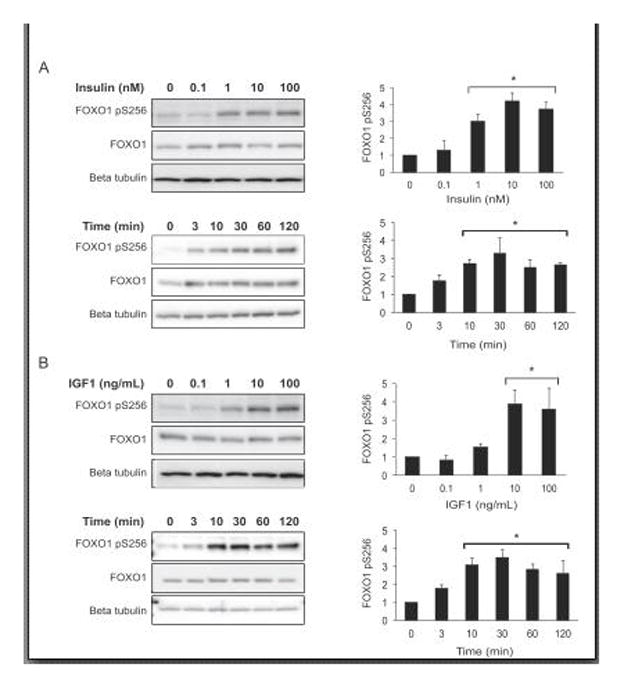
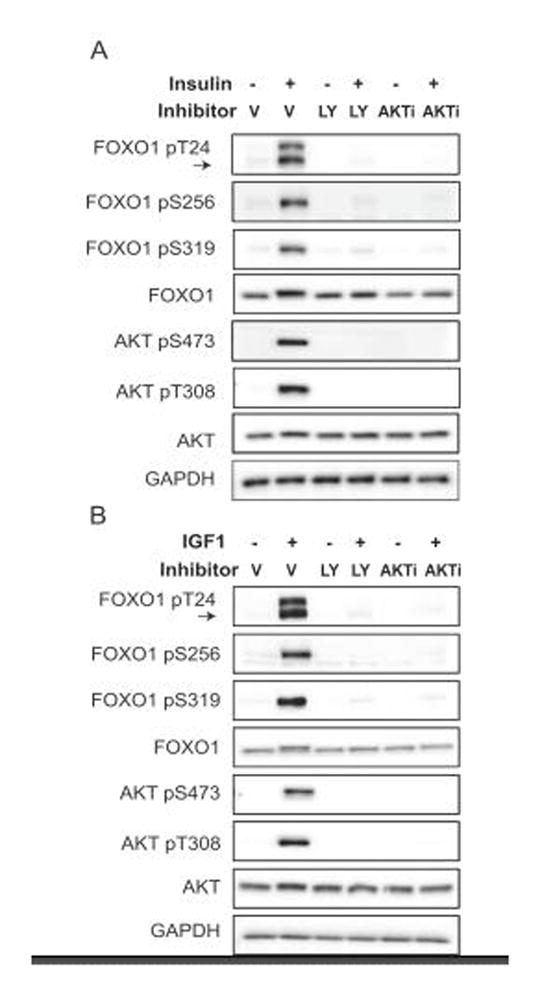
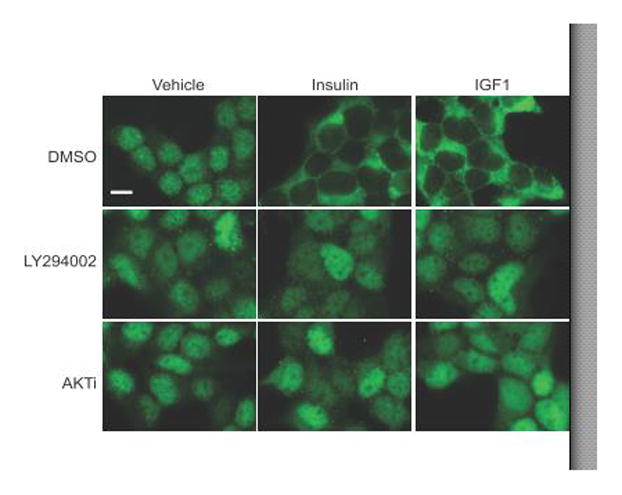
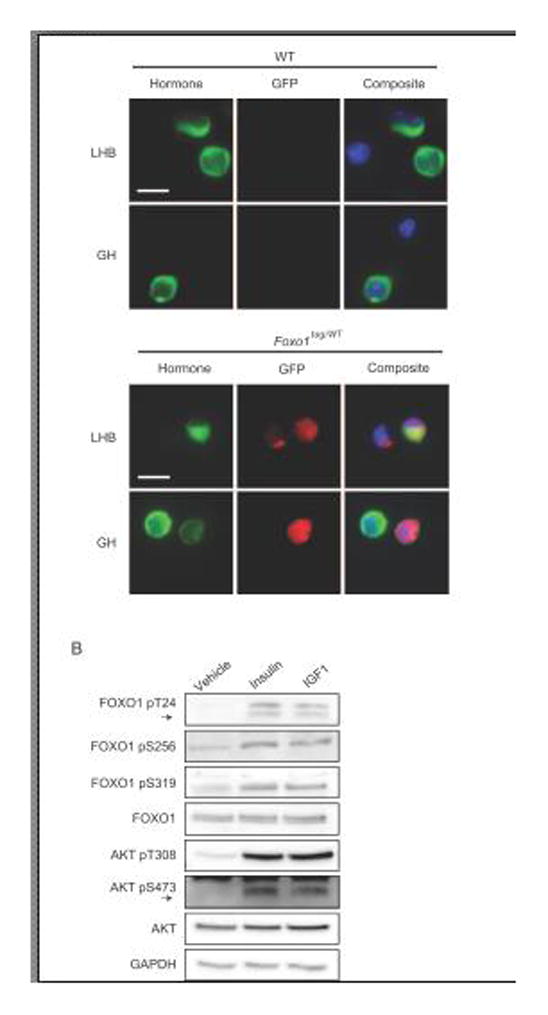
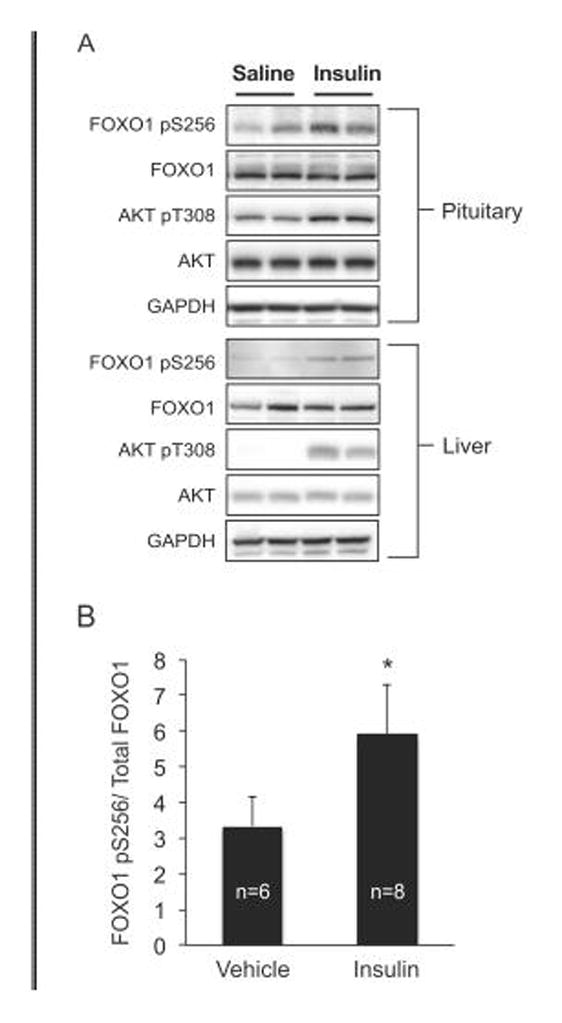
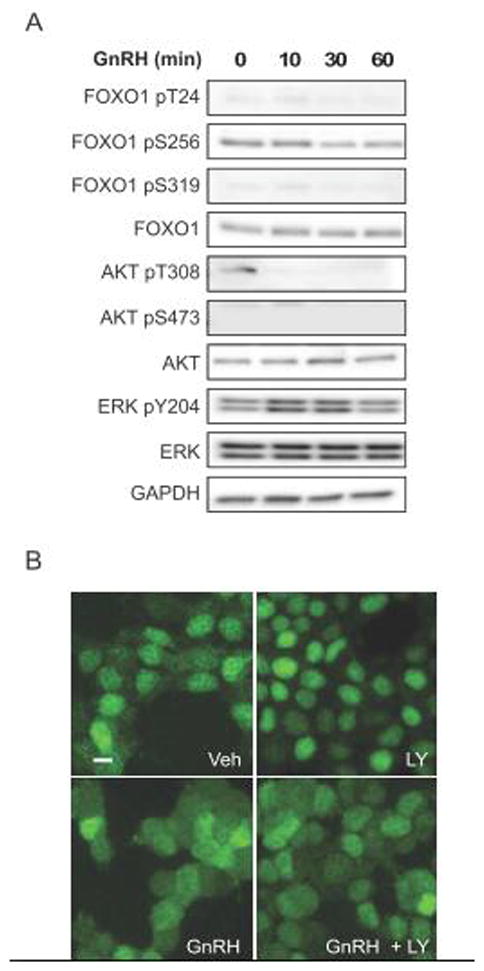
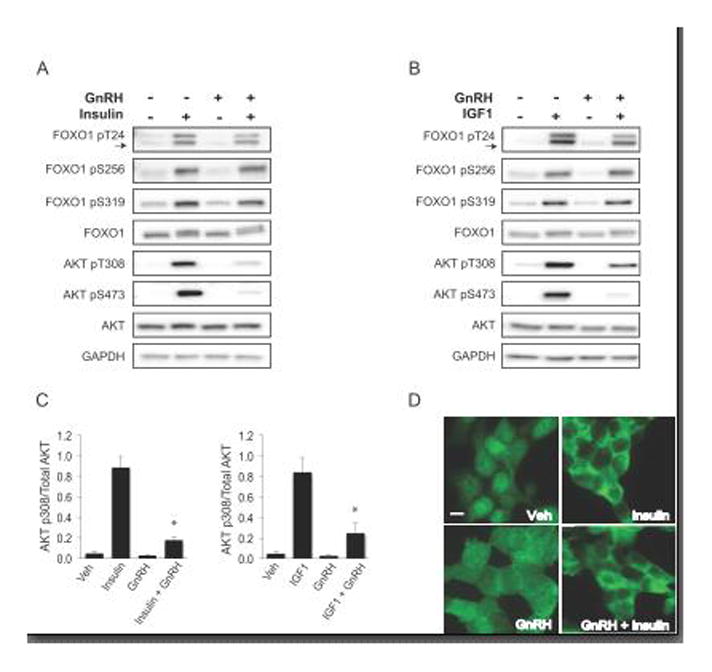
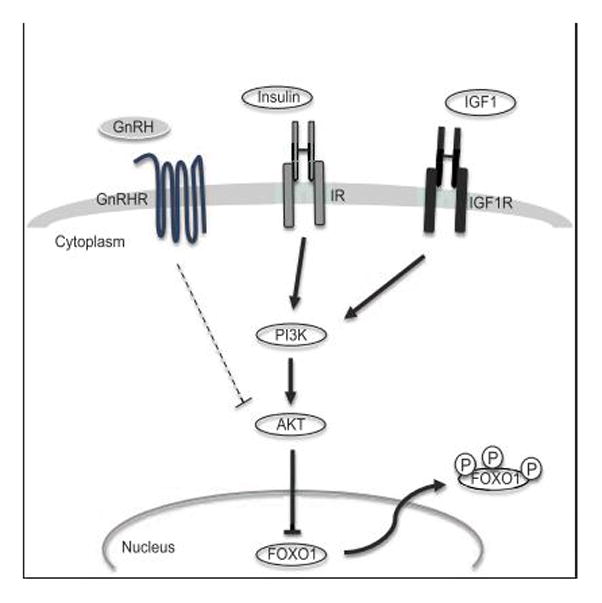
The FOXO1 transcription factor is important for multiple aspects of reproductive function. We previously reported that FOXO1 functions as a repressor of gonadotropin hormone synthesis, but how FOXO1 is regulated in pituitary gonadotropes is unknown. The growth factors, insulin and insulin-like growth factor I (IGF1), function as key regulators of cell proliferation, metabolism and apoptosis in multiple cell types through the PI3K/AKT signaling pathway. In this study, we found that insulin and IGF1 signaling in gonadotropes induced FOXO1 phosphorylation through the PI3K/AKT pathway in immortalized and primary cells, resulting in FOXO1 relocation from the nucleus to the cytoplasm. Furthermore, insulin administration in vivo induced phosphorylation of FOXO1 and AKT in the pituitary. Thus, insulin and IGF1 act as negative regulators of FOXO1 activity and may serve to fine-tune gonadotropin expression.
Keywords: Forkhead box O1; GnRH; Gonadotrope; IGF1; Insulin.
Copyright © 2015 Elsevier Ireland Ltd. All rights reserved.
Figures
References
-
- Galili N, Davis RJ, Fredericks WJ, Mukhopadhyay S, Rauscher FJ, 3rd, Emanuel BS, Rovera G, Barr FG. Fusion of a fork head domain gene to PAX3 in the solid tumour alveolar rhabdomyosarcoma. Nat Genet. 1993;5:230–235. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
