

Functions of the 5' and 3' ends of calicivirus genomes
- PMID: 25678268
- PMCID: PMC4509552
- DOI: 10.1016/j.virusres.2015.02.002
Functions of the 5' and 3' ends of calicivirus genomes
Abstract
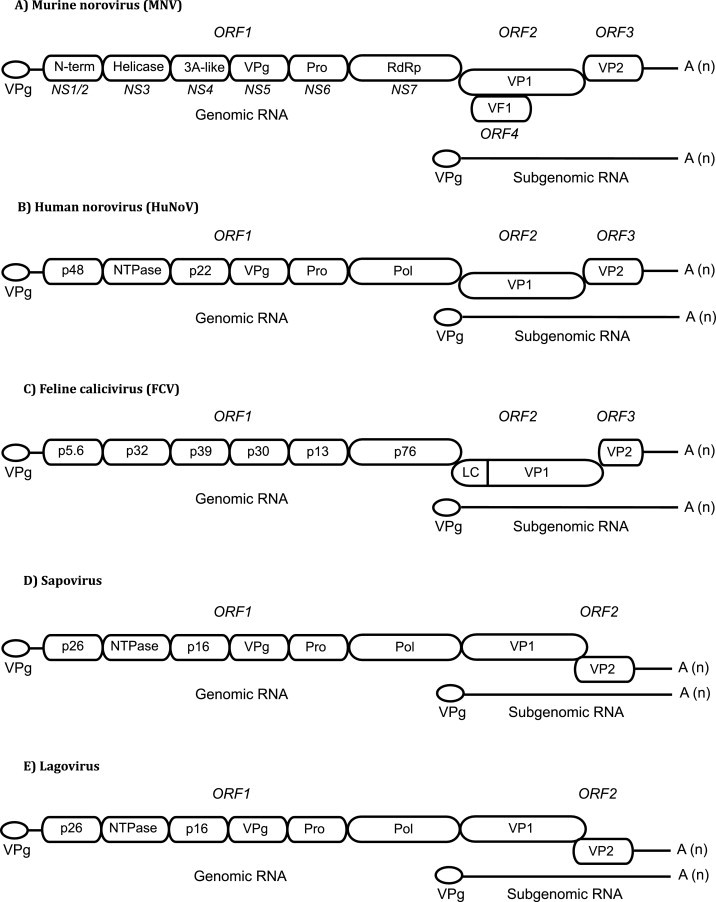
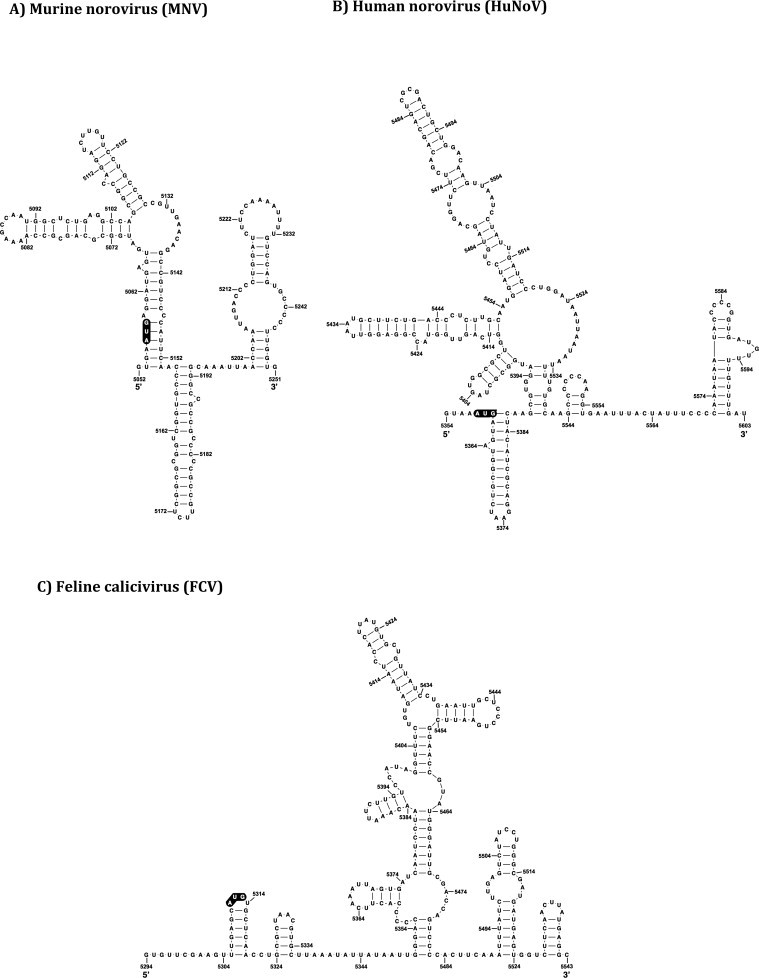
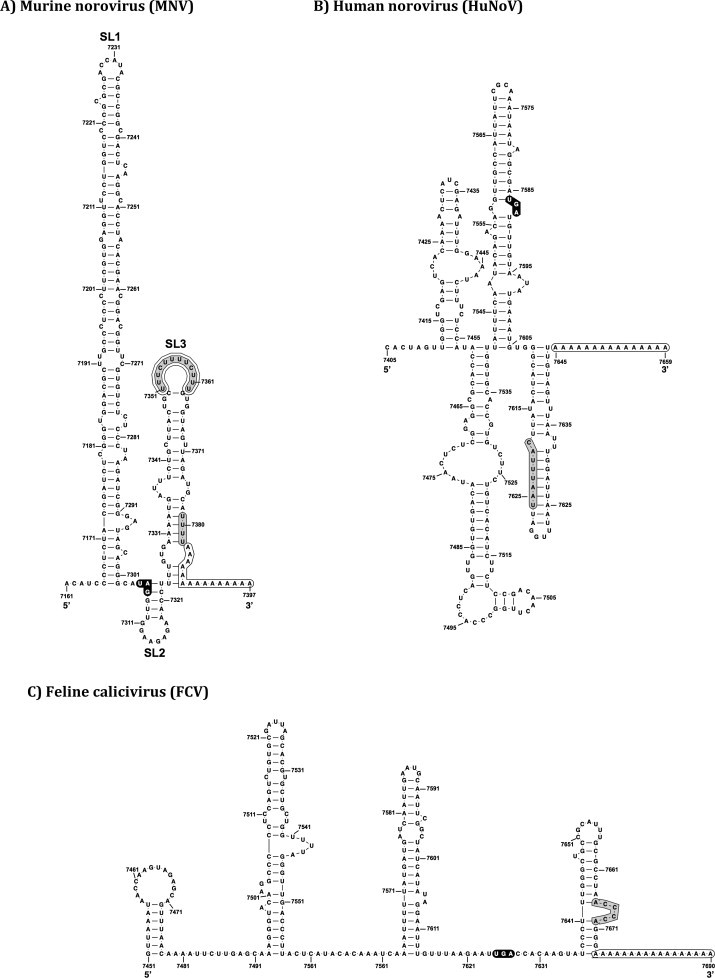
The Caliciviridae family of small positive sense RNA viruses contains a diverse range of pathogens of both man and animals. The molecular mechanisms of calicivirus genome replication and translation have not been as widely studied as many other RNA viruses. With the relatively recent development of robust cell culture and reverse genetics systems for several members of the Caliciviridae family, a more in-depth analysis of the finer detail of the viral life cycle has now been obtained. As a result, the identification and characterization of the role of RNA structures in the calicivirus life cycle has also been possible. This review aims to summarize the current state of knowledge with respect to the role of RNA structures at the termini of calicivirus genomes.
Keywords: Caliciviruses; Norovirus; RNA secondary structures; RNA–protein interactions.
Copyright © 2015 The Authors. Published by Elsevier B.V. All rights reserved.
Figures

References
-
- Agis-Juárez R.A., Galván I., Medina F., Daikoku T., Padmanabhan R., Ludert J.E., del Angel R.M. Polypyrimidine tract-binding protein is relocated to the cytoplasm and is required during dengue virus infection in Vero cells. J. Gen. Virol. 2009;90:2893–2901. doi: 10.1099/vir.0.013433-0. - DOI - PubMed
-
- Bailey D., Karakasiliotis I., Vashist S., Chung L.M.W., Rees J., Reese J., McFadden N., Benson A., Yarovinsky F., Simmonds P., Goodfellow I. Functional analysis of RNA structures present at the 3′ extremity of the murine norovirus genome: the variable polypyrimidine tract plays a role in viral virulence. J. Virol. 2010;84:2859–2870. doi: 10.1128/JVI.02053-09. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
