

Structural chromosome abnormalities, increased DNA strand breaks and DNA strand break repair deficiency in dermal fibroblasts from old female human donors
- PMID: 25678531
- PMCID: PMC4359693
- DOI: 10.18632/aging.100723
Structural chromosome abnormalities, increased DNA strand breaks and DNA strand break repair deficiency in dermal fibroblasts from old female human donors
Abstract
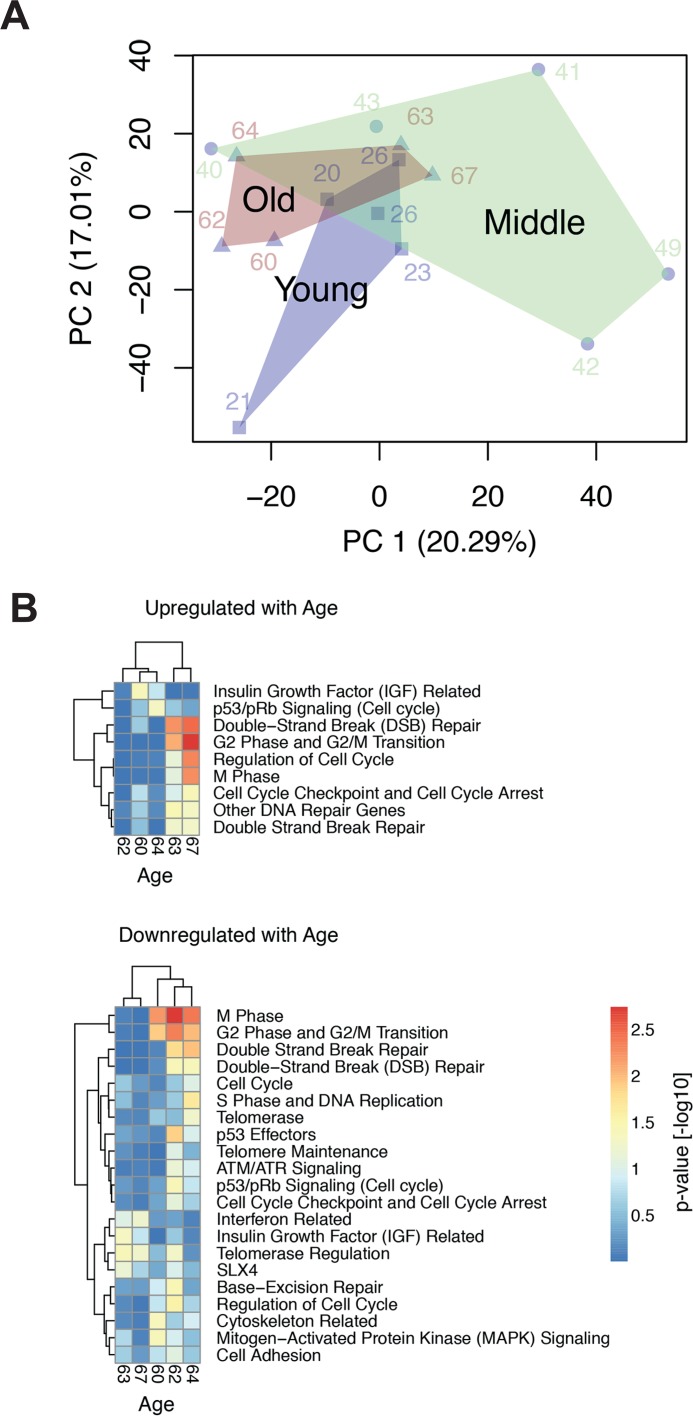
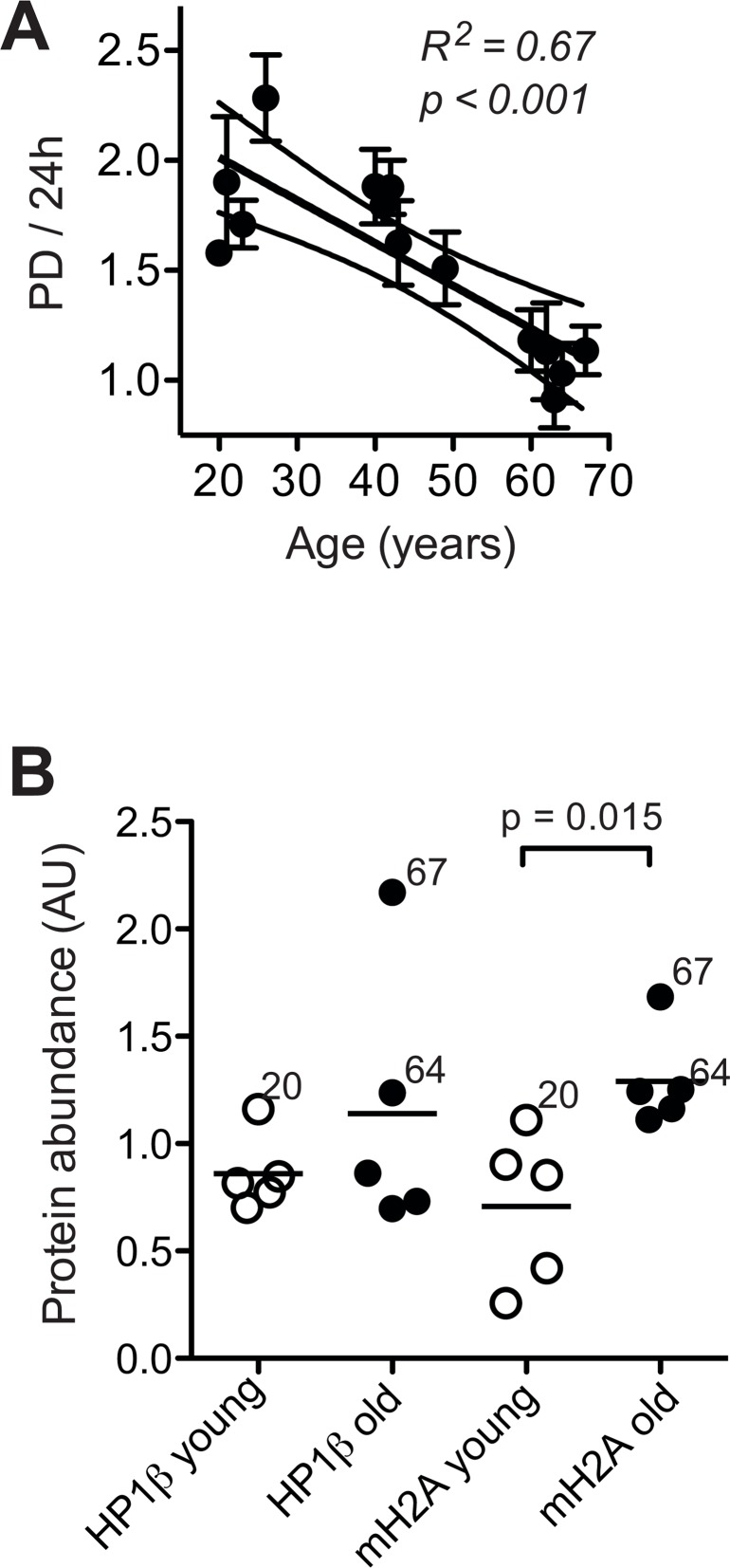
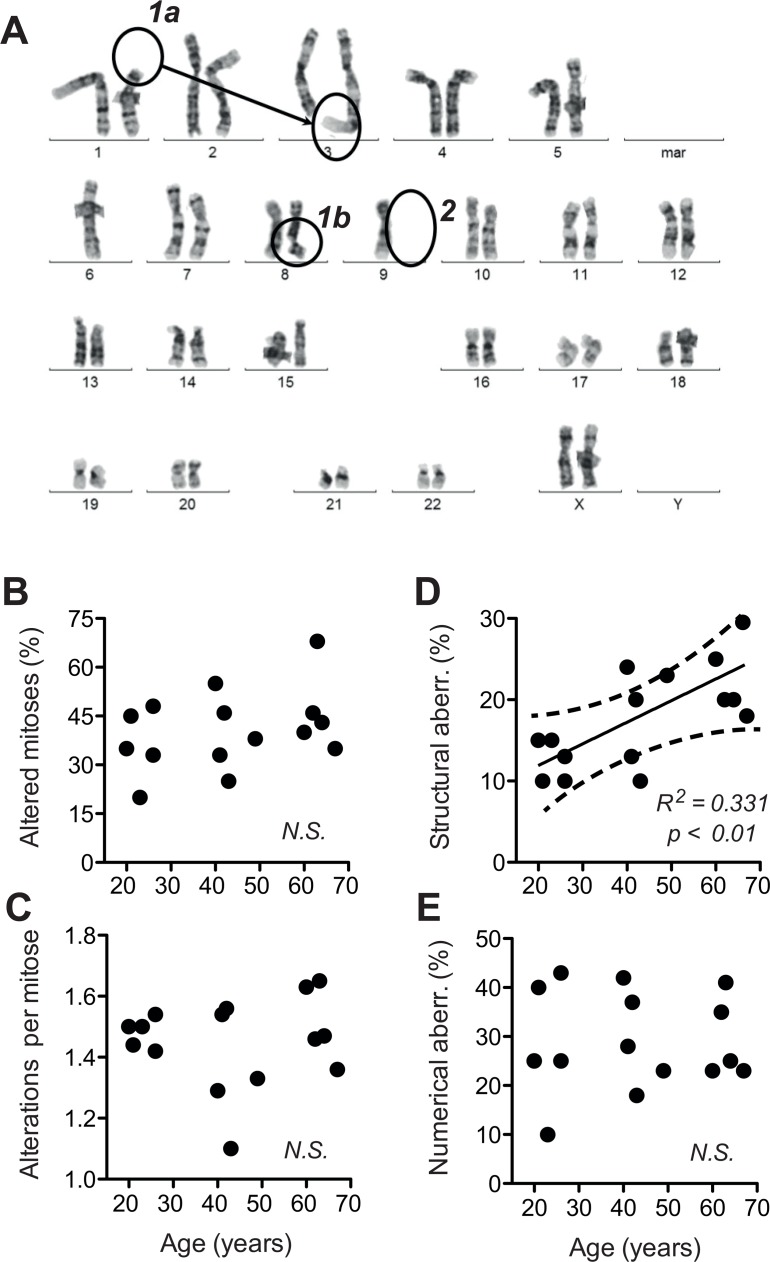
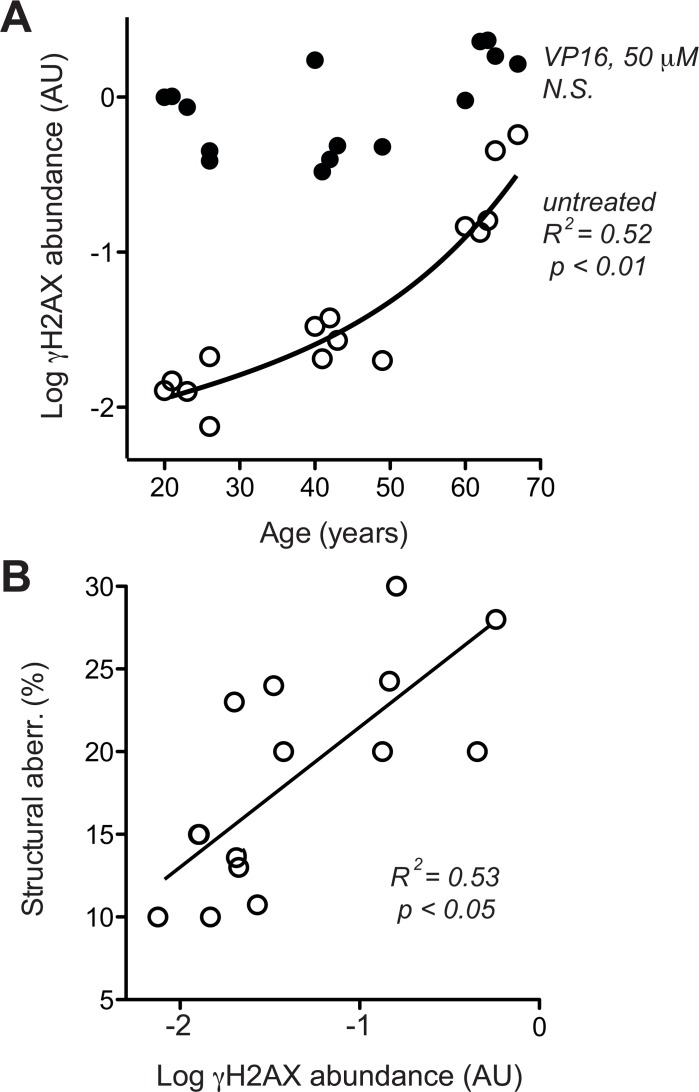
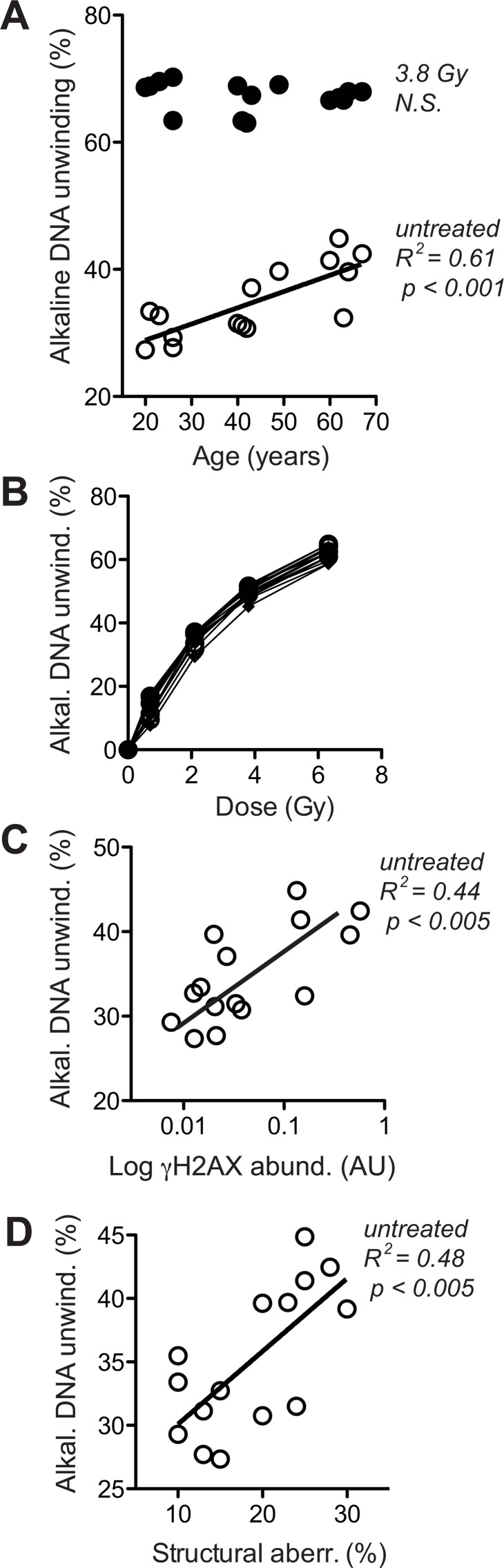
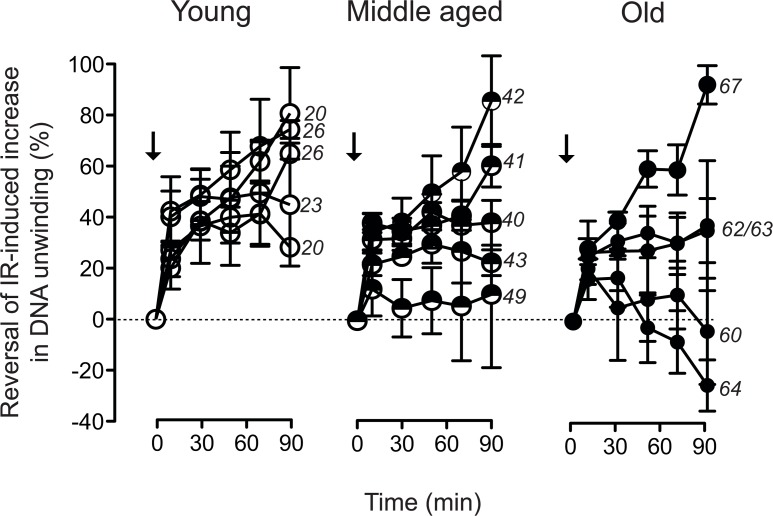
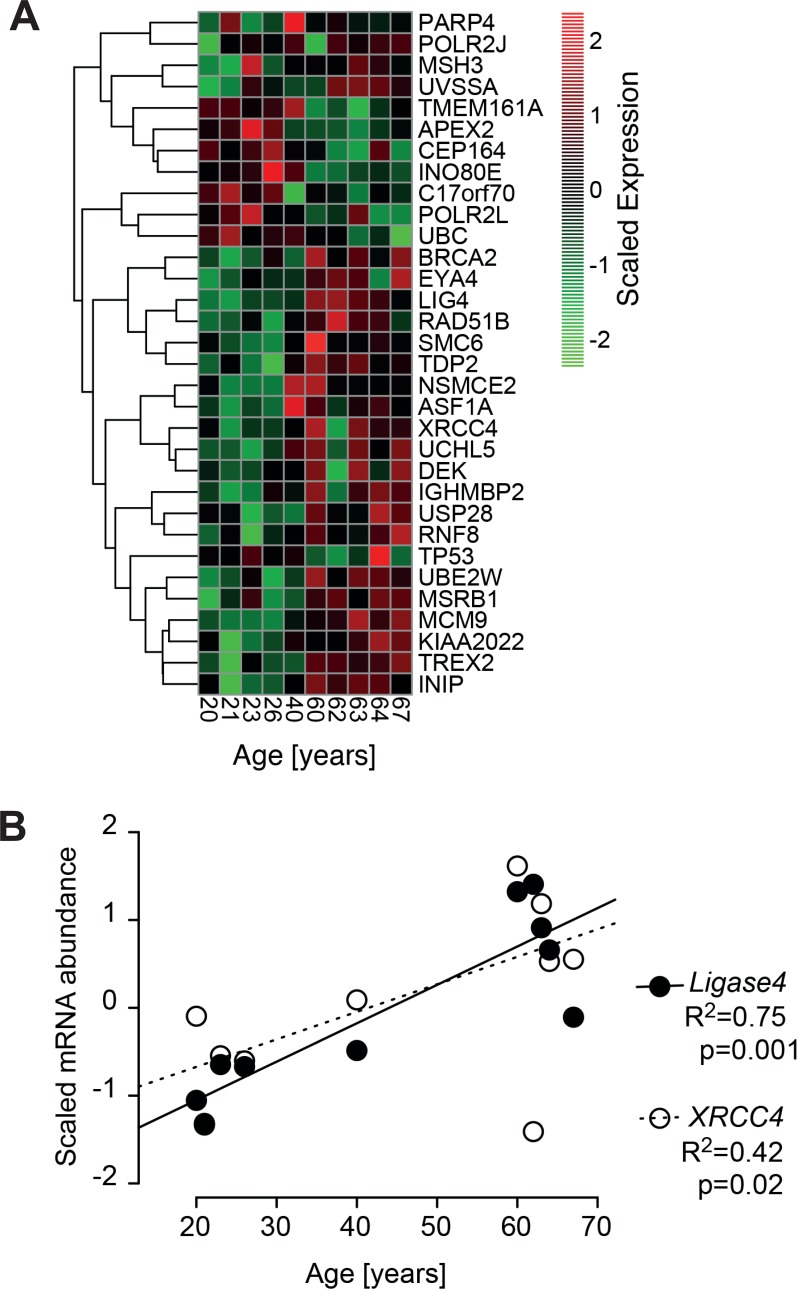
Dermal fibroblasts provide a paradigmatic model of cellular adaptation to long-term exogenous stress and ageing processes driven thereby. Here we addressed whether fibroblast ageing analysedex vivo entails genome instability. Dermal fibroblasts from human female donors aged 20-67 years were studied in primary culture at low population doubling. Under these conditions, the incidence of replicative senescence and rates of age-correlated telomere shortening were insignificant. Genome-wide gene expression analysis revealed age-related impairment of mitosis, telomere and chromosome maintenance and induction of genes associated with DNA repair and non-homologous end-joining, most notably XRCC4 and ligase 4. We observed an age-correlated drop in proliferative capacity and age-correlated increases in heterochromatin marks, structural chromosome abnormalities (deletions, translocations and chromatid breaks), DNA strand breaks and histone H2AX-phosphorylation. In a third of the cells from old and middle-aged donors repair of X-ray induced DNA strand breaks was impaired despite up-regulation of DNA repair genes. The distinct phenotype of genome instability, increased heterochromatinisation and (in 30% of the cases futile) up-regulation of DNA repair genes was stably maintained over several cell passages indicating that it represents a feature of geroconversion that is distinct from cellular senescence, as it does not encompass a block of proliferation.
Conflict of interest statement
The authors have no conflict of interests to declare.
Figures
References
-
- Boukamp P. Skin aging: a role for telomerase and telomere dynamics? Curr Mol Med. 2005;5:171–177. - PubMed
-
- Tigges J, Krutmann J, Fritsche E, Haendeler J, Schaal H, Fischer JW, Kalfalah F, Reinke H, Reifenberger G, Stuhler K, Ventura N, Gundermann S, Boukamp P, et al. The hallmarks of fibroblast ageing. Mech Ageing Dev. 2014;138:26–44. - PubMed
-
- Schulze C, Wetzel F, Kueper T, Malsen A, Muhr G, Jaspers S, Blatt T, Wittern KP, Wenck H, Kas JA. Stiffening of human skin fibroblasts with age. Clin Plast Surg. 2012;39:9–20. - PubMed
-
- Boraldi F, Bini L, Liberatori S, Armini A, Pallini V, Tiozzo R, Pasquali-Ronchetti I, Quaglino D. Proteome analysis of dermal fibroblasts cultured in vitro from human healthy subjects of different ages. Proteomics. 2003;3:917–929. - PubMed
-
- Kueper T, Grune T, Prahl S, Lenz H, Welge V, Biernoth T, Vogt Y, Muhr GM, Gaemlich A, Jung T, Boemke G, Elsasser HP, Wittern KP, et al. Vimentin is the specific target in skin glycation. Structural prerequisites, functional consequences, and role in skin aging. J Biol Chem. 2007;282:23427–23436. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
