

Modulation of nociceptive dural input to the trigeminocervical complex through GluK1 kainate receptors
- PMID: 25679470
- PMCID: PMC7611087
- DOI: 10.1097/01.j.pain.0000460325.25762.c0
Modulation of nociceptive dural input to the trigeminocervical complex through GluK1 kainate receptors
Abstract
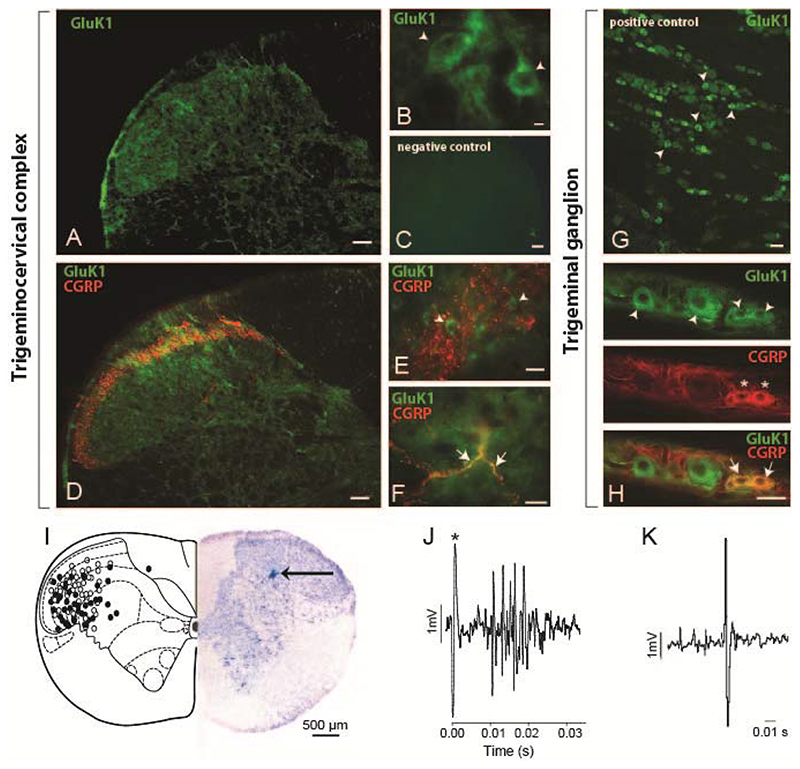
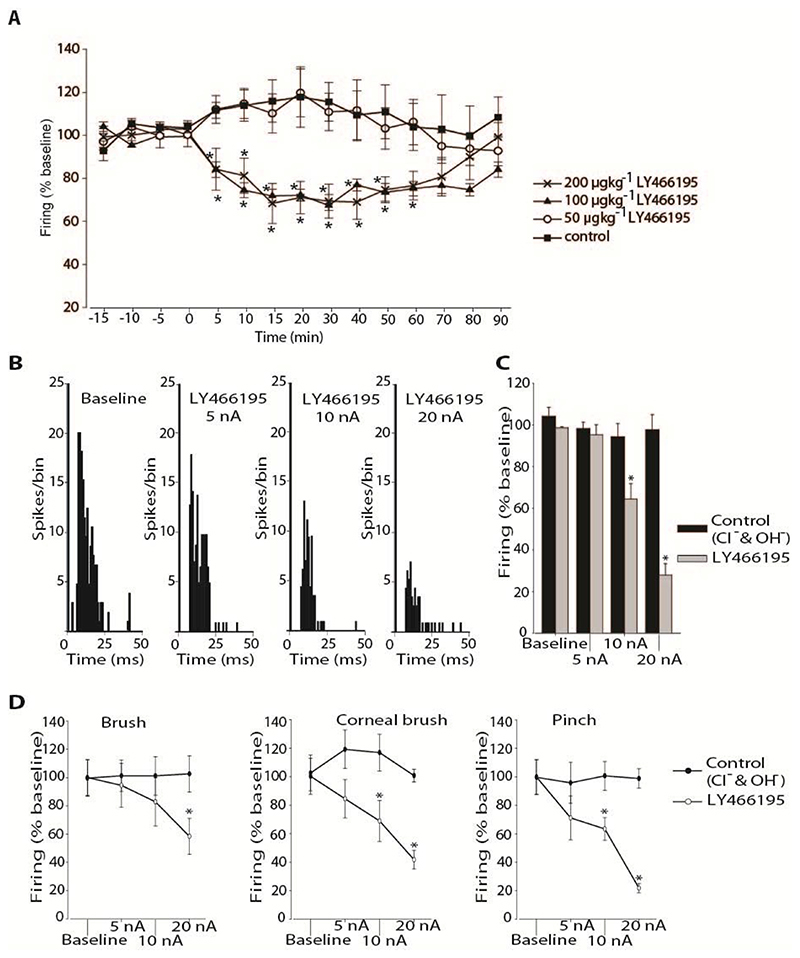
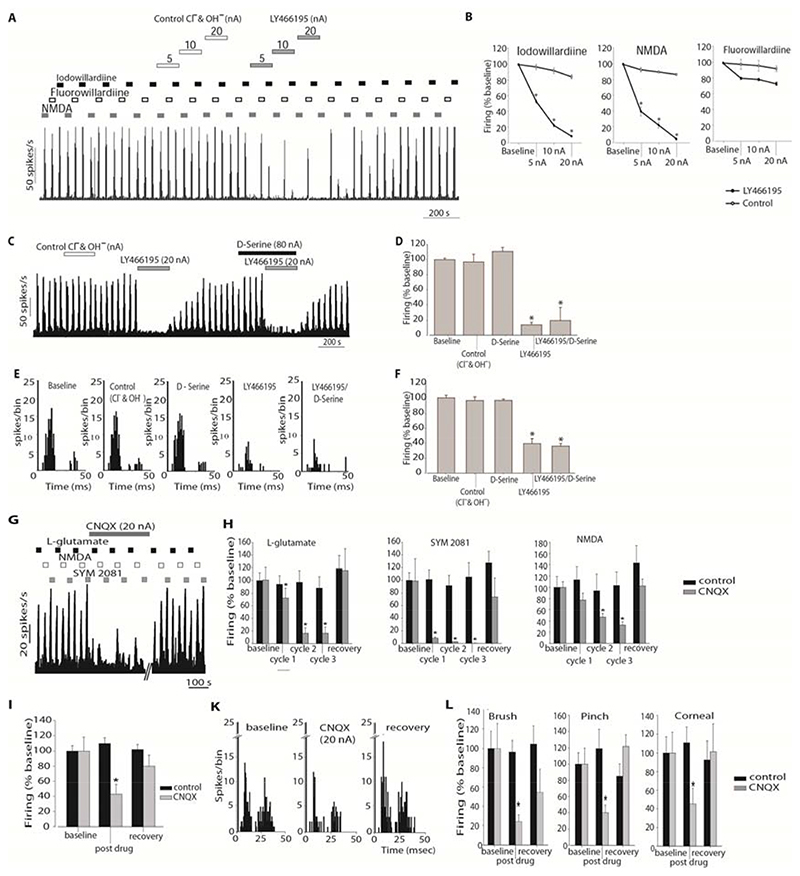
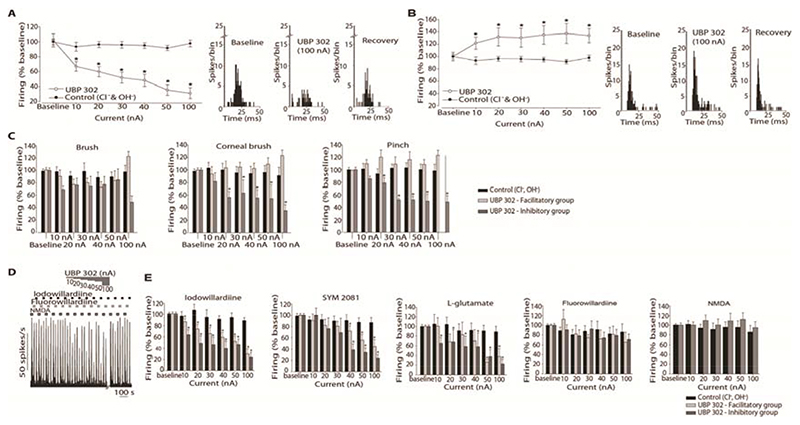
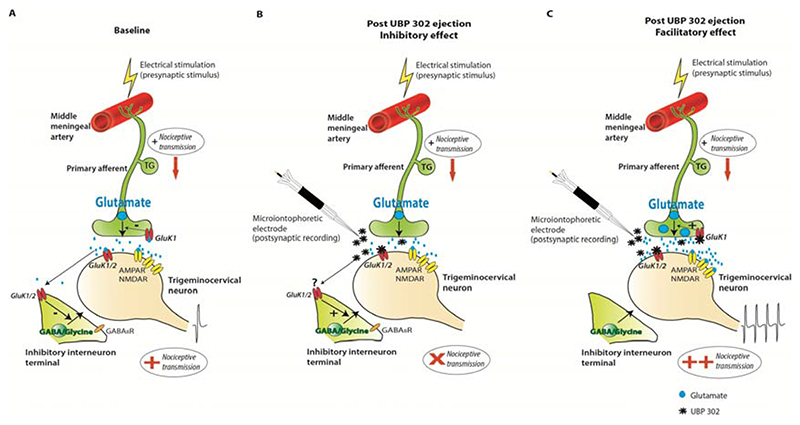
Migraine is a common and disabling neurologic disorder, with important psychiatric comorbidities. Its pathophysiology involves activation of neurons in the trigeminocervical complex (TCC). Kainate receptors carrying the glutamate receptor subunit 5 (GluK1) are present in key brain areas involved in migraine pathophysiology. To study the influence of kainate receptors on trigeminovascular neurotransmission, we determined the presence of GluK1 receptors within the trigeminal ganglion and TCC with immunohistochemistry. We performed in vivo electrophysiologic recordings from TCC neurons and investigated whether local or systemic application of GluK1 receptor antagonists modulated trigeminovascular transmission. Microiontophoretic application of a selective GluK1 receptor antagonist, but not of a nonspecific ionotropic glutamate receptor antagonist, markedly attenuated cell firing in a subpopulation of neurons activated in response to dural stimulation, consistent with selective inhibition of postsynaptic GluK1 receptor-evoked firing seen in all recorded neurons. In contrast, trigeminovascular activation was significantly facilitated in a different neuronal population. The clinically active kainate receptor antagonist LY466195 attenuated trigeminovascular activation in all neurons. In addition, LY466195 demonstrated an N-methyl-d-aspartate receptor-mediated effect. This study demonstrates a differential role of GluK1 receptors in the TCC, antagonism of which can inhibit trigeminovascular activation through postsynaptic mechanisms. Furthermore, the data suggest a novel, possibly presynaptic, modulatory role of trigeminocervical kainate receptors in vivo. Differential activation of kainate receptors suggests unique roles for this receptor in pro- and antinociceptive mechanisms in migraine pathophysiology.
Conflict of interest statement
The other authors declare no competing financial interests.
Figures
References
-
- Afridi S, Giffin NJ, Kaube H, Goadsby PJ. A randomized controlled trial of intranasal ketamine in migraine with prolonged aura. Neurology. 2013;80:642–647. - PubMed
-
- Akerman S, Goadsby PJ. Topiramate inhibits cortical spreading depression in rat and cat: impact in migraine aura. Neuroreport. 2005;16(12):1383–1387. - PubMed
-
- Andreou AP, Goadsby PJ. Therapeutic potential of novel glutamate receptor antagonists in migraine. Expert opinion on investigational drugs. 2009;18(6):789–803. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
