

Targeting the adaptability of heterogeneous aneuploids
- PMID: 25679766
- PMCID: PMC4328141
- DOI: 10.1016/j.cell.2015.01.026
Targeting the adaptability of heterogeneous aneuploids
Abstract
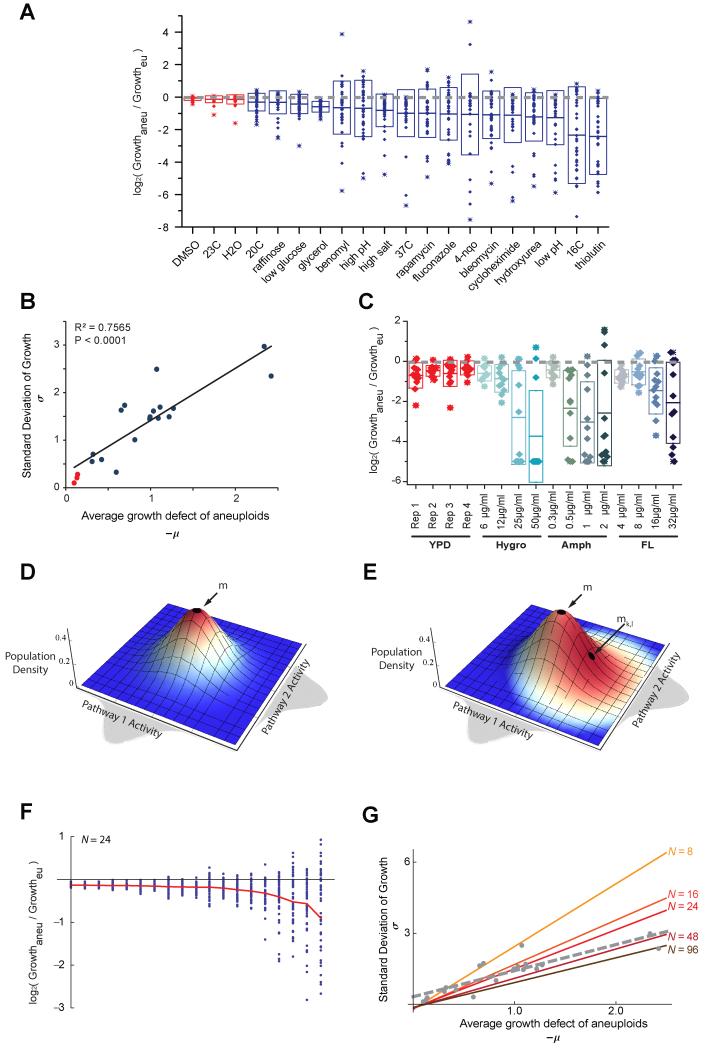
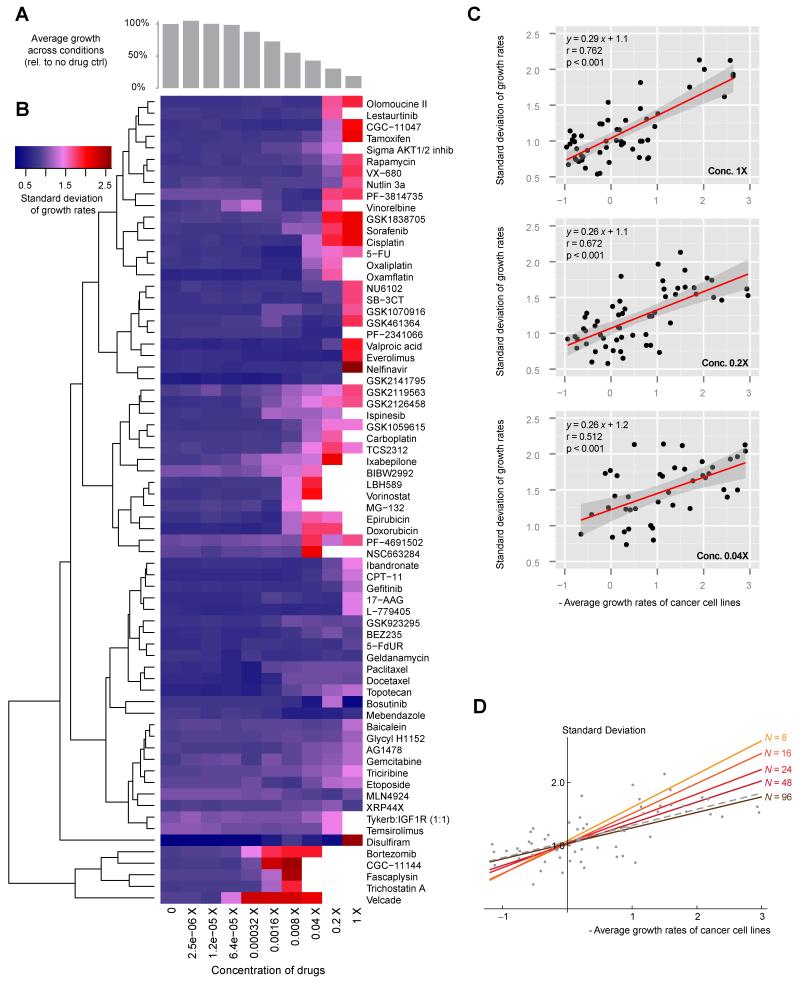
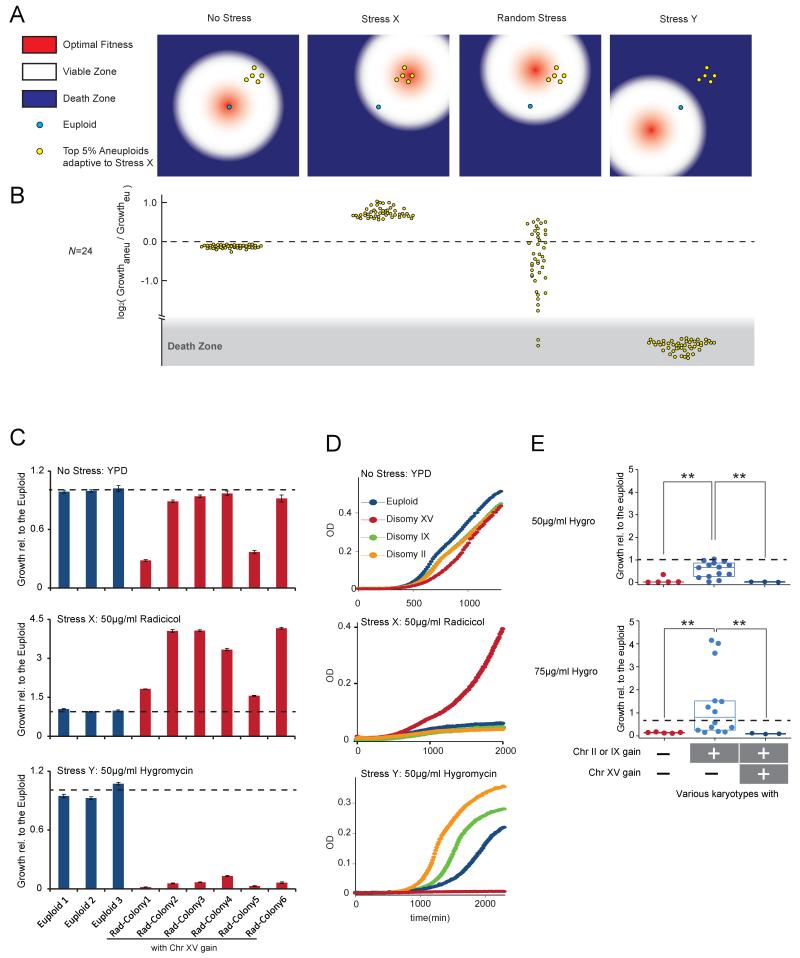
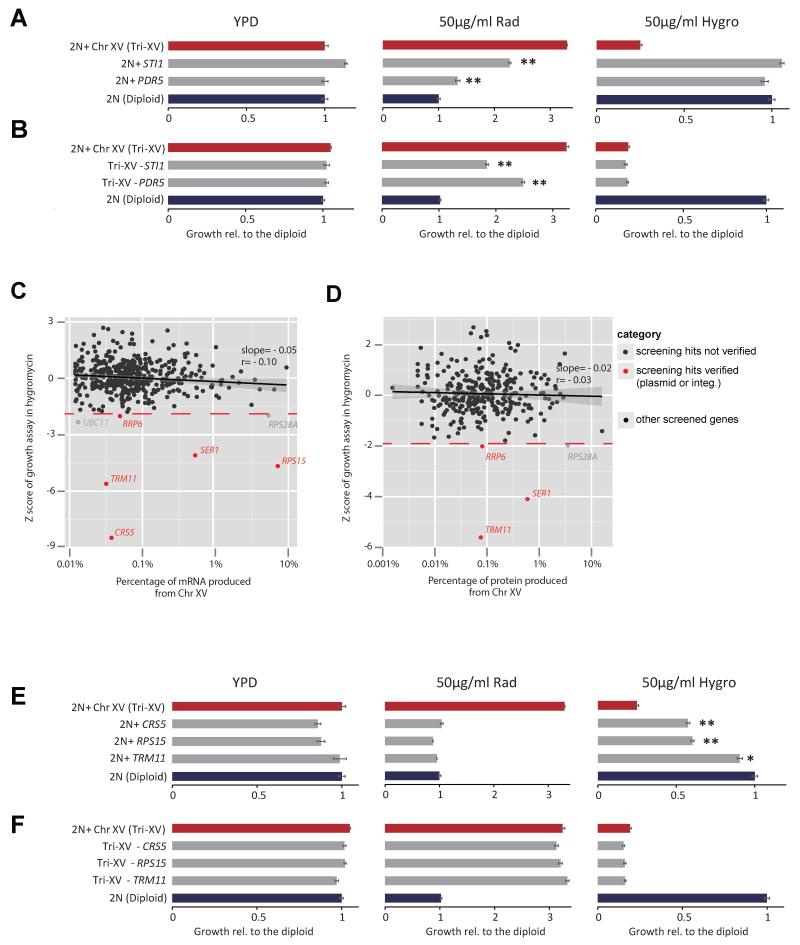
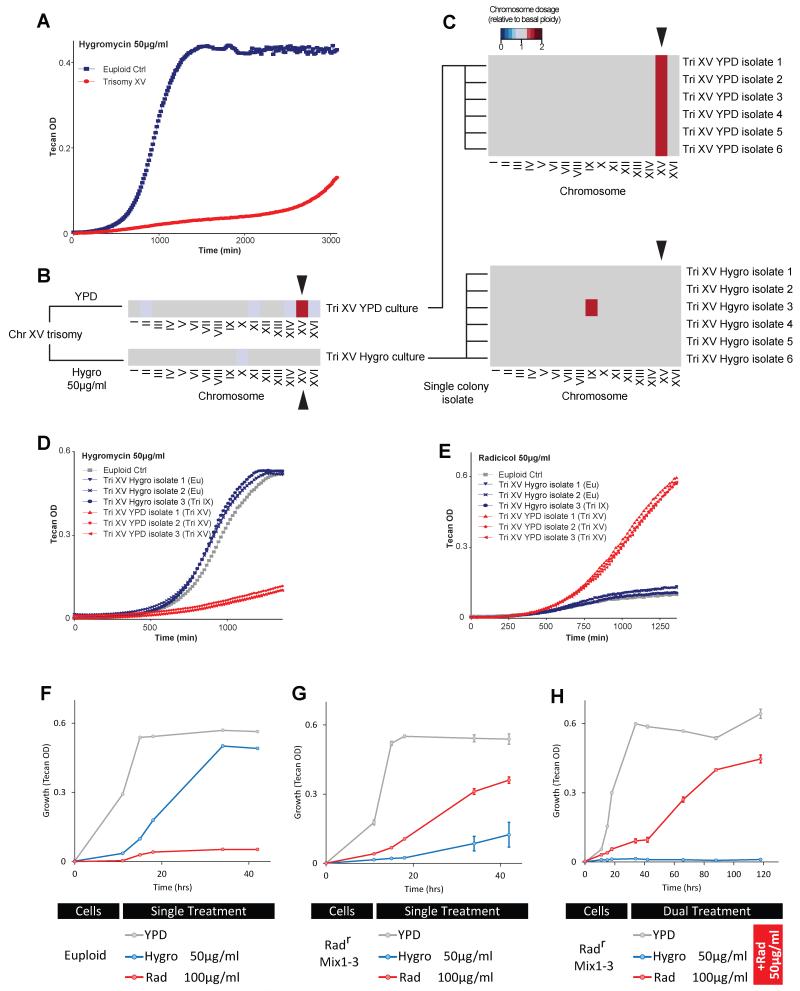
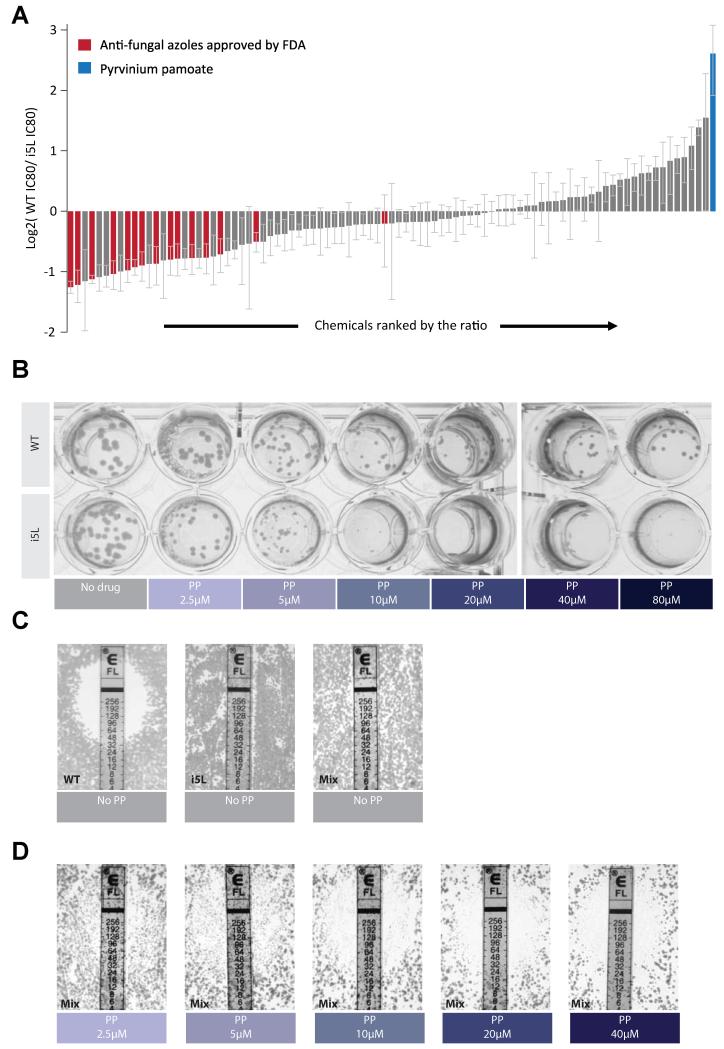
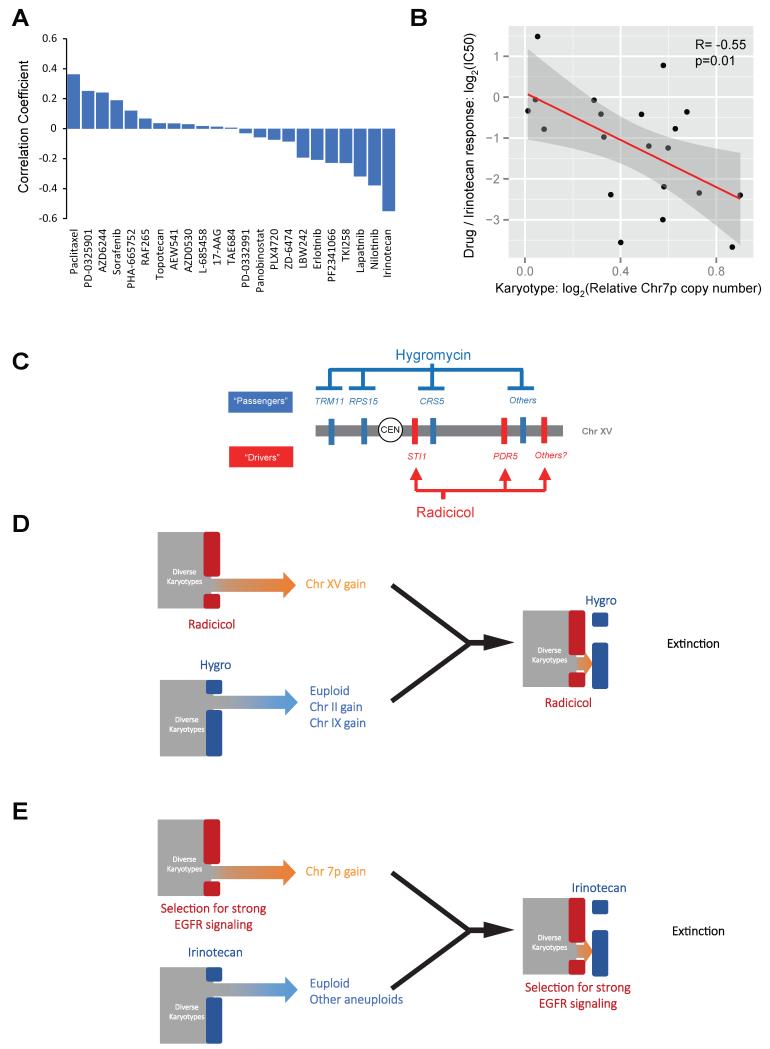
Aneuploid genomes, characterized by unbalanced chromosome stoichiometry (karyotype), are associated with cancer malignancy and drug resistance of pathogenic fungi. The phenotypic diversity resulting from karyotypic diversity endows the cell population with superior adaptability. We show here, using a combination of experimental data and a general stochastic model, that the degree of phenotypic variation, thus evolvability, escalates with the degree of overall growth suppression. Such scaling likely explains the challenge of treating aneuploidy diseases with a single stress-inducing agent. Instead, we propose the design of an "evolutionary trap" (ET) targeting both karyotypic diversity and fitness. This strategy entails a selective condition "channeling" a karyotypically divergent population into one with a predominant and predictably drugable karyotypic feature. We provide a proof-of-principle case in budding yeast and demonstrate the potential efficacy of this strategy toward aneuploidy-based azole resistance in Candida albicans. By analyzing existing pharmacogenomics data, we propose the potential design of an ET against glioblastoma.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Bean J, Brennan C, Shih J-Y, Riely G, Viale A, Wang L, Chitale D, Motoi N, Szoke J, Broderick S, et al. MET amplification occurs with or without T790M mutations in EGFR mutant lung tumors with acquired resistance to gefitinib or erlotinib. Proceedings of the National Academy of Sciences. 2007;104:20932–20937. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
