

Functional differences in the acinar cells of the murine major salivary glands
- PMID: 25680367
- PMCID: PMC4502782
- DOI: 10.1177/0022034515570943
Functional differences in the acinar cells of the murine major salivary glands
Abstract
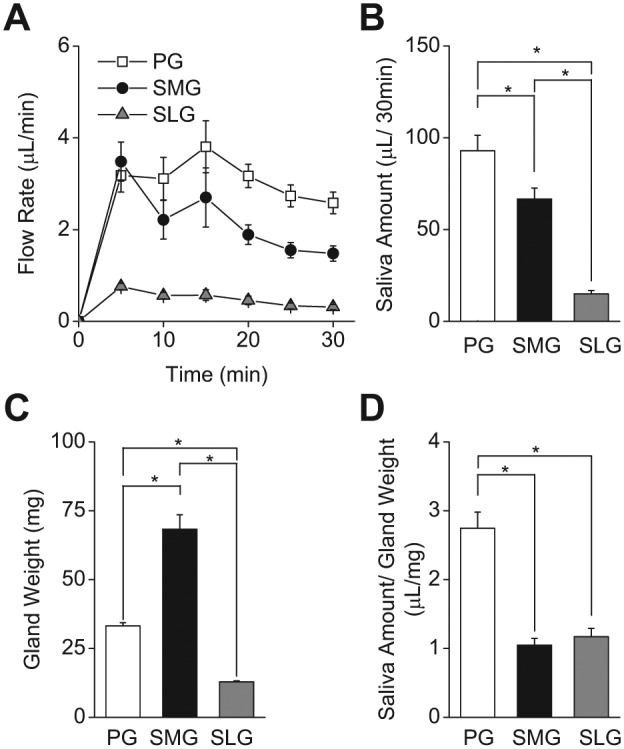
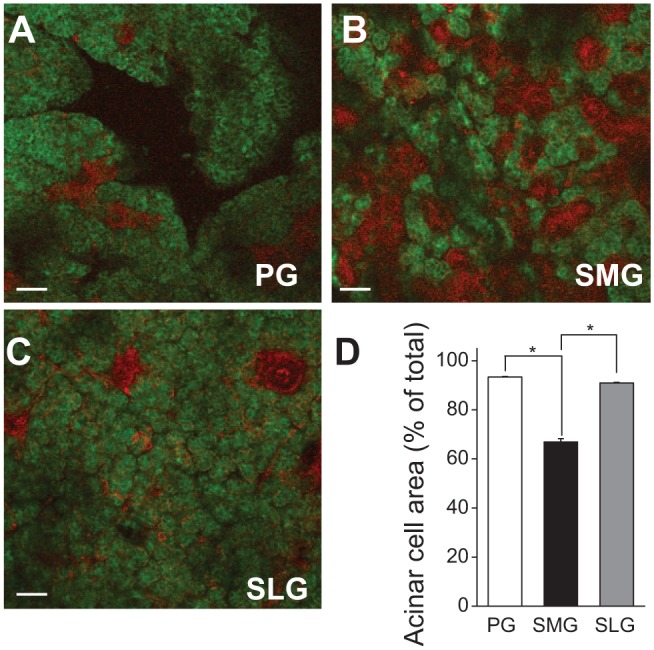
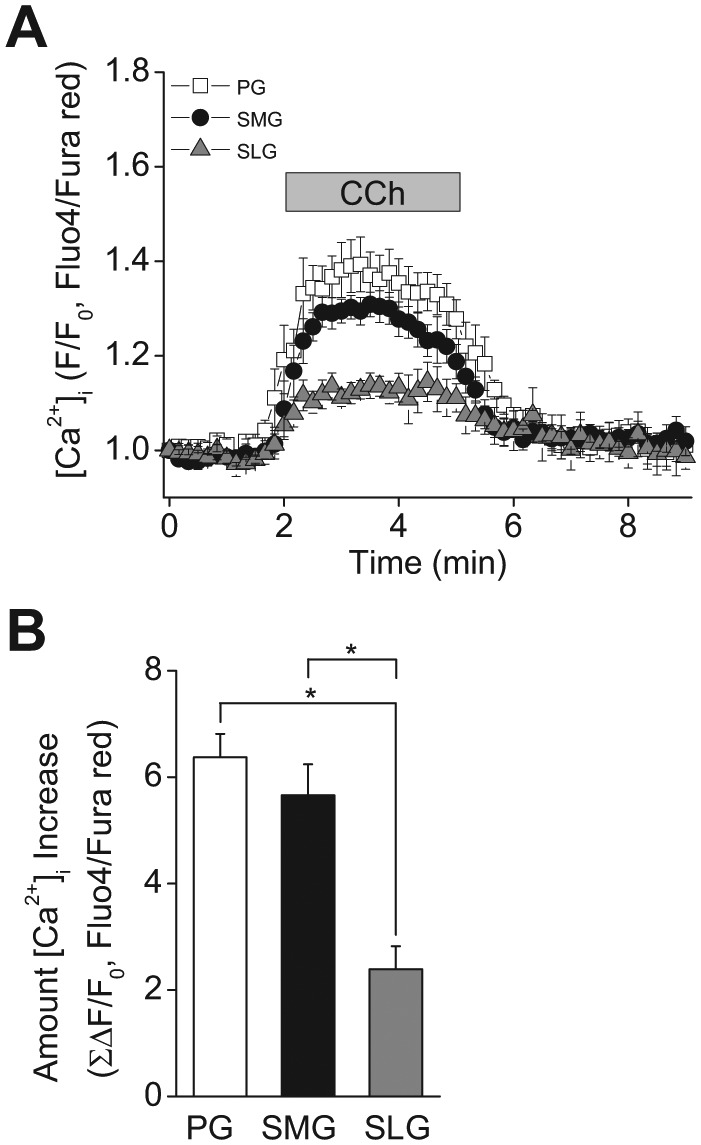
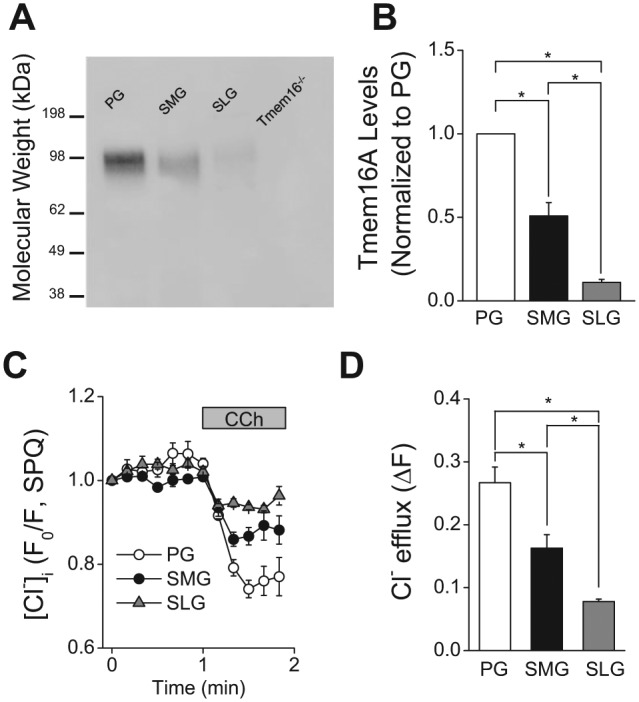
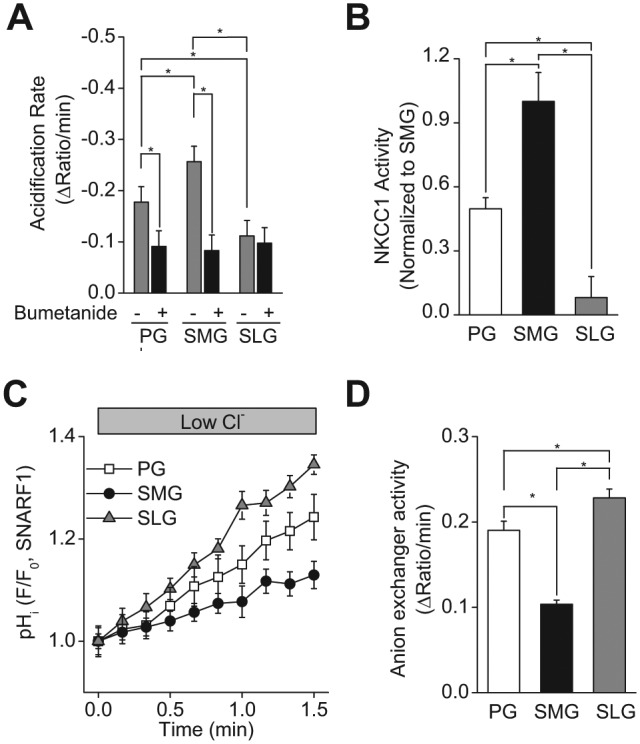
In humans, approximately 90% of saliva is secreted by the 3 major salivary glands: the parotid (PG), the submandibular (SMG), and the sublingual glands (SLG). Even though it is known that all 3 major salivary glands secrete saliva by a Cl(-)-dependent mechanism, salivary secretion rates differ greatly among these glands. The goal of this study was to gain insight into the properties of the ion-transporting pathways in acinar cells that might account for the differences among the major salivary glands. Pilocarpine-induced saliva was simultaneously collected in vivo from the 3 major salivary glands of mice. When normalized by gland weight, the amount of saliva secreted by the PG was more than 2-fold larger than that obtained from the SMG and SLG. At the cellular level, carbachol induced an increase in the intracellular [Ca(2+)] that was more than 2-fold larger in PG and SMG than in SLG acinar cells. Carbachol-stimulated Cl(-) efflux and the protein levels of the Ca(2+)-activated Cl(-) channel TMEM16A, the major apical Cl(-) efflux pathway in salivary acinar cells, were significantly greater in PG compared with SMG and SLG. In addition, we evaluated the transporter activity of the Na(+)-K(+)-2Cl(-) cotransporters (NKCC1) and anion exchangers (AE), the 2 primary basolateral Cl(-) uptake mechanisms in acinar cells. The SMG NKCC1 activity was about twice that of the PG and more than 12-fold greater than that of the SLG. AE activity was similar in PG and SLG, and both PG and SLG AE activity was about 2-fold larger than that of SMG. In summary, the salivation kinetics of the 3 major glands are distinct, and these differences can be explained by the unique functional properties of each gland related to Cl(-) movement, including the transporter activities of the Cl(-) uptake and efflux pathways, and intracellular Ca(2+) mobilization.
Keywords: Na-K-Cl transporter; calcium signaling; chloride channel; epithelia; physiology; salivary physiology.
© International & American Associations for Dental Research 2015.
Conflict of interest statement
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
Figures
References
-
- Caputo A, Caci E, Ferrera L, Pedemonte N, Barsanti C, Sondo E, Pfeffer U, Ravazzolo R, Zegarra-Moran O, Galietta LJ. 2008. TMEM16A, a membrane protein associated with calcium-dependent chloride channel activity. Science. 322(5901):590–594. - PubMed
-
- Dawes C, Wood CM. 1973. The contribution of oral minor mucous gland secretions to the volume of whole saliva in man. Arch Oral Biol. 18(3):337–342. - PubMed
-
- Edgar WM. 1990. Saliva and dental health. Clinical implications of saliva: report of a consensus meeting. Br Dent J. 169(3-4):96–98. - PubMed
-
- Evans RL, Park K, Turner RJ, Watson GE, Nguyen HV, Dennett MR, Hand AR, Flagella M, Shull GE, Melvin JE. 2000. Severe impairment of salivation in Na+/K+/2Cl- cotransporter (NKCC1)-deficient mice. J Biol Chem. 275(35):26720–26726. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
