

Arabidopsis glutamate receptor homolog3.5 modulates cytosolic Ca2+ level to counteract effect of abscisic acid in seed germination
- PMID: 25681329
- PMCID: PMC4378146
- DOI: 10.1104/pp.114.251298
Arabidopsis glutamate receptor homolog3.5 modulates cytosolic Ca2+ level to counteract effect of abscisic acid in seed germination
Abstract
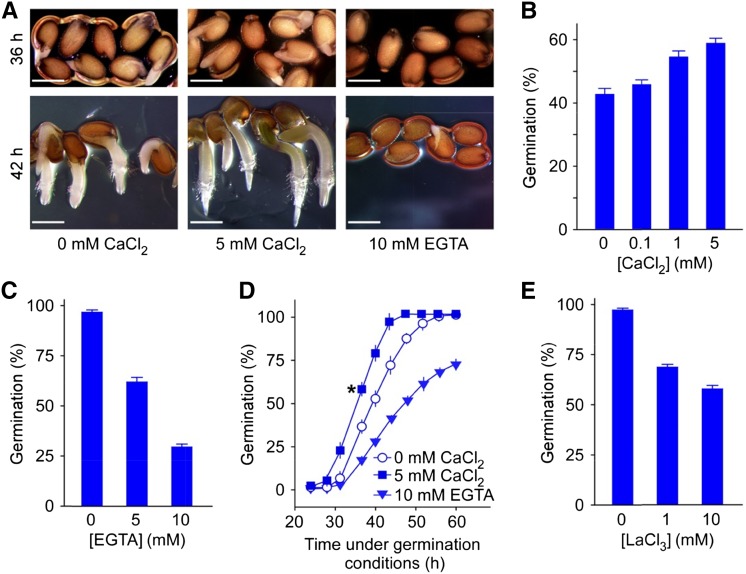
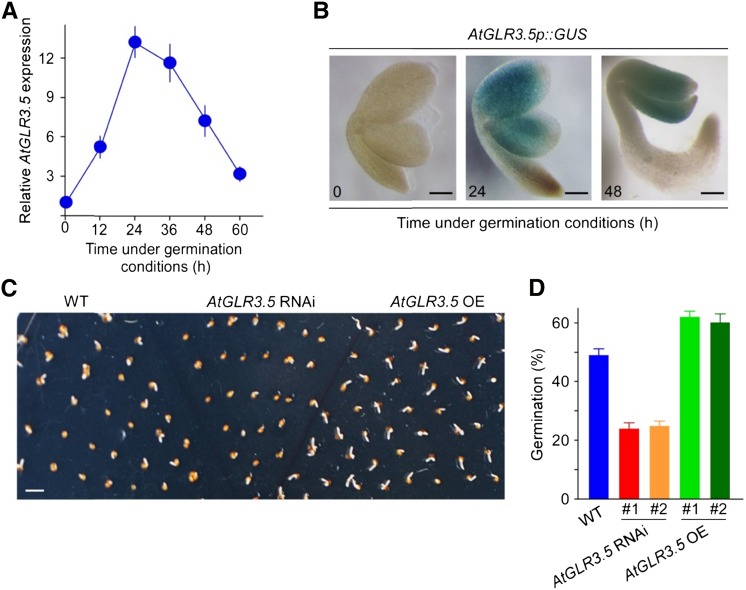
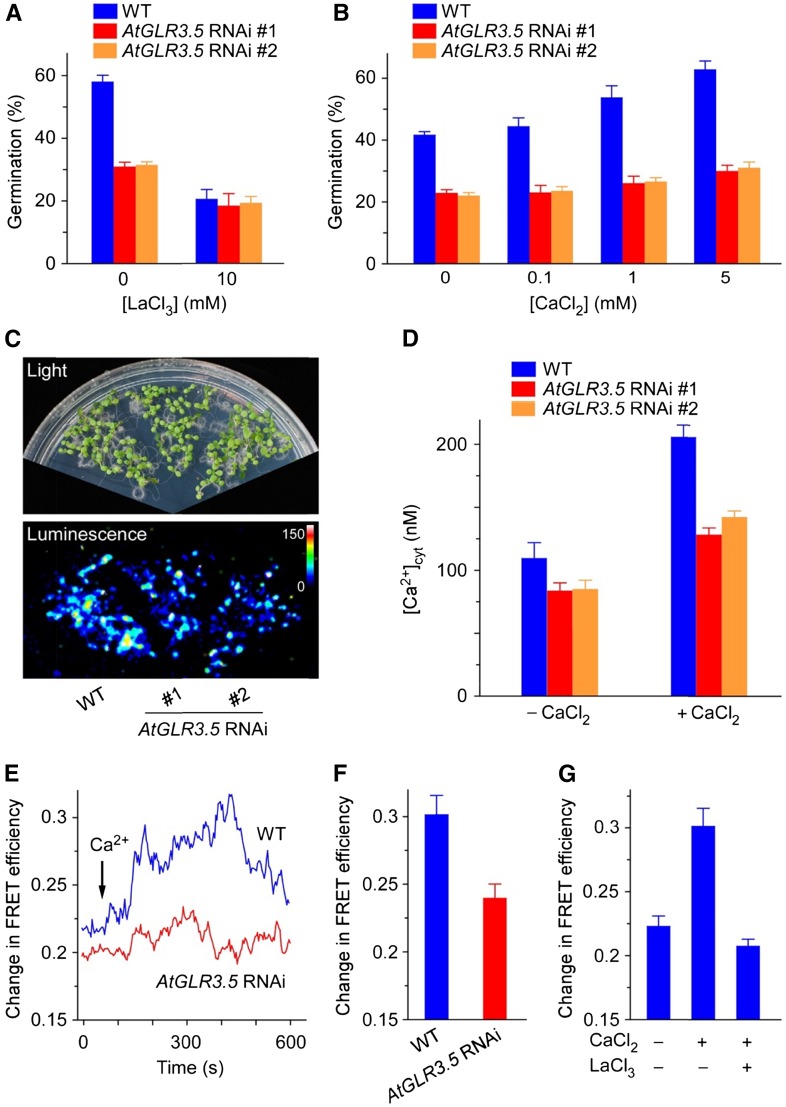
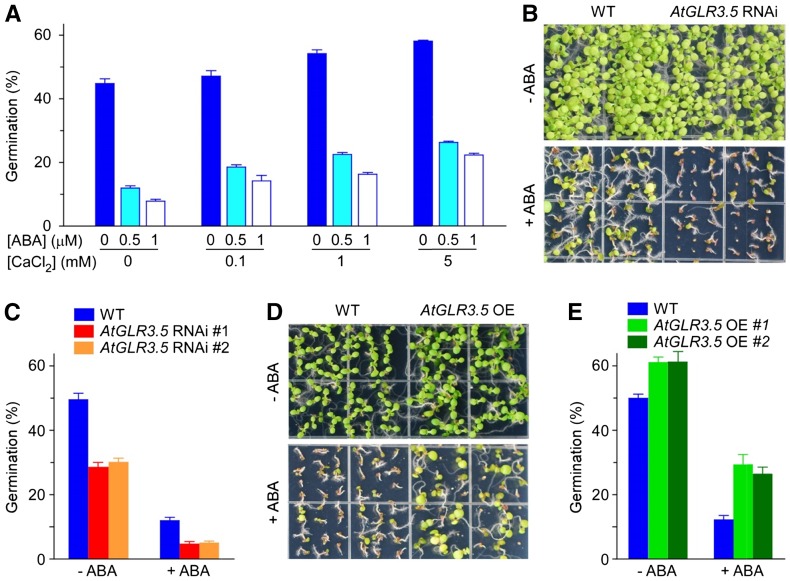
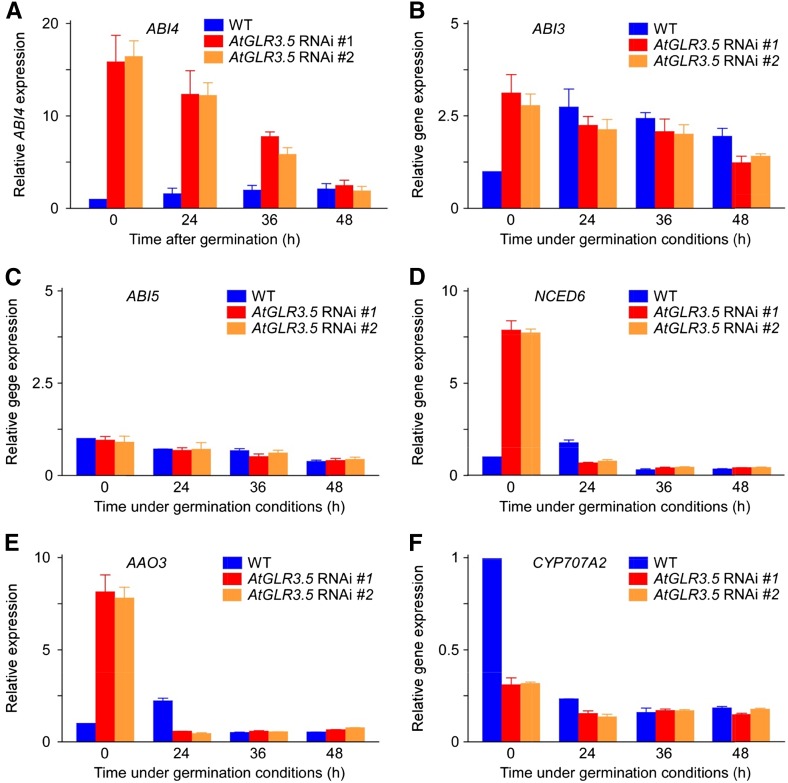
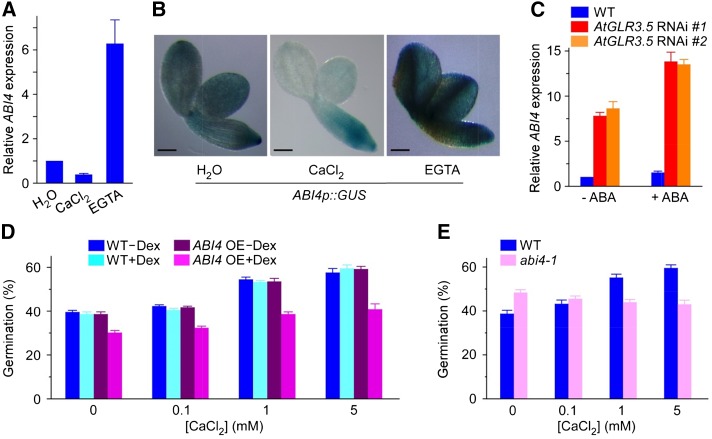
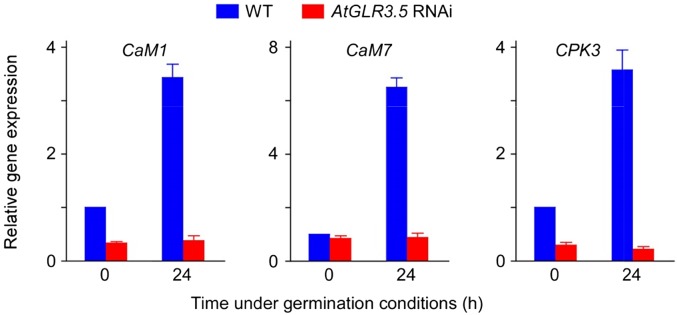
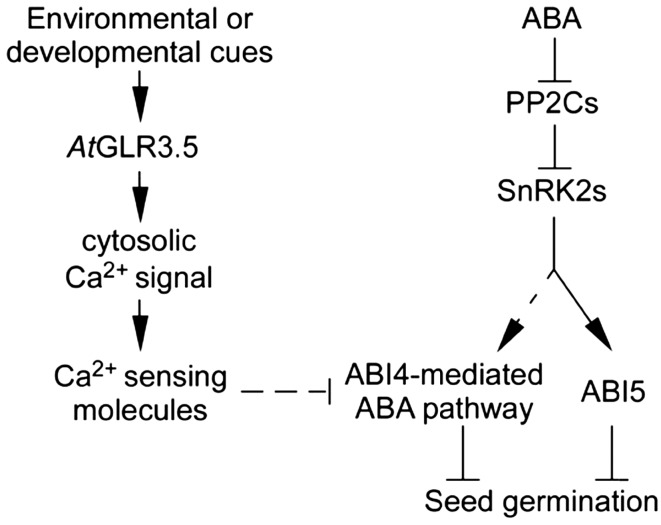
Seed germination is a critical step in a plant's life cycle that allows successful propagation and is therefore strictly controlled by endogenous and environmental signals. However, the molecular mechanisms underlying germination control remain elusive. Here, we report that the Arabidopsis (Arabidopsis thaliana) glutamate receptor homolog3.5 (AtGLR3.5) is predominantly expressed in germinating seeds and increases cytosolic Ca2+ concentration that counteracts the effect of abscisic acid (ABA) to promote germination. Repression of AtGLR3.5 impairs cytosolic Ca2+ concentration elevation, significantly delays germination, and enhances ABA sensitivity in seeds, whereas overexpression of AtGLR3.5 results in earlier germination and reduced seed sensitivity to ABA. Furthermore, we show that Ca2+ suppresses the expression of ABSCISIC ACID INSENSITIVE4 (ABI4), a key transcription factor involved in ABA response in seeds, and that ABI4 plays a fundamental role in modulation of Ca2+-dependent germination. Taken together, our results provide molecular genetic evidence that AtGLR3.5-mediated Ca2+ influx stimulates seed germination by antagonizing the inhibitory effects of ABA through suppression of ABI4. These findings establish, to our knowledge, a new and pivotal role of the plant glutamate receptor homolog and Ca2+ signaling in germination control and uncover the orchestrated modulation of the AtGLR3.5-mediated Ca2+ signal and ABA signaling via ABI4 to fine-tune the crucial developmental process, germination, in Arabidopsis.
© 2015 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Bailly C, El-Maarouf-Bouteau H, Corbineau F (2008) From intracellular signaling networks to cell death: the dual role of reactive oxygen species in seed physiology. C R Biol 331: 806–814 - PubMed
-
- Bassel GW, Mullen RT, Bewley JD (2006) ABI3 expression ceases following, but not during, germination of tomato and Arabidopsis seeds. J Exp Bot 57: 1291–1297 - PubMed
-
- Bonilla I, El-Hamdaoui A, Bolanos L (2004) Boron and calcium increase Pisum sativum seed germination and seedling development under salt stress. Plant Soil 267: 97–107
-
- Chiu JC, Brenner ED, DeSalle R, Nitabach MN, Holmes TC, Coruzzi GM (2002) Phylogenetic and expression analysis of the glutamate-receptor-like gene family in Arabidopsis thaliana. Mol Biol Evol 19: 1066–1082 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
