

Dynamics of cross-bridge cycling, ATP hydrolysis, force generation, and deformation in cardiac muscle
- PMID: 25681584
- PMCID: PMC4532654
- DOI: 10.1016/j.yjmcc.2015.02.006
Dynamics of cross-bridge cycling, ATP hydrolysis, force generation, and deformation in cardiac muscle
Abstract
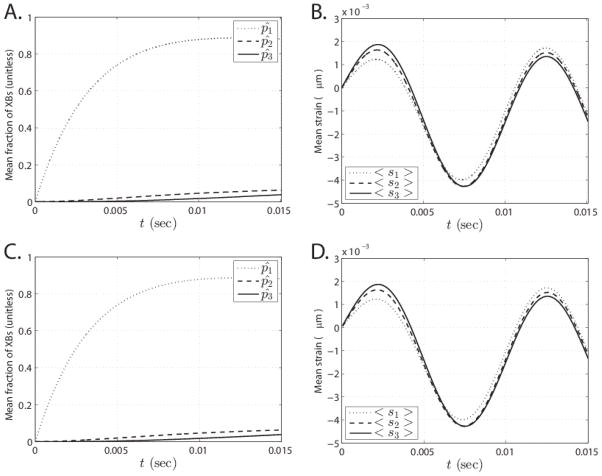
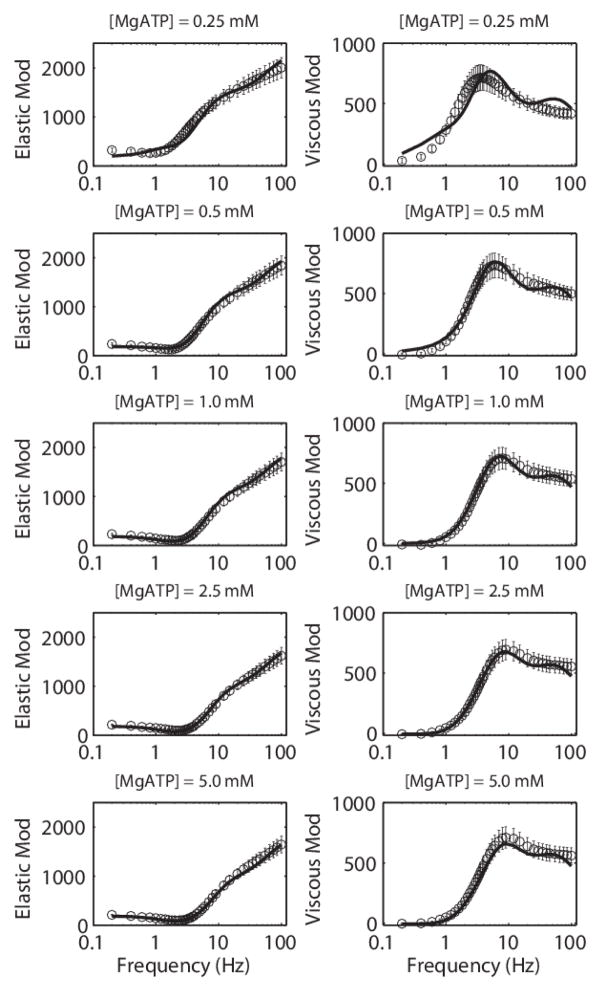
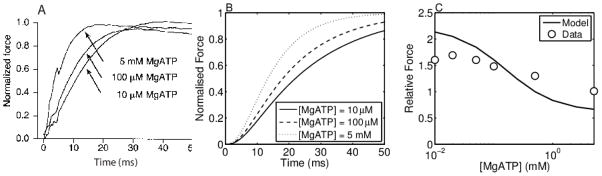
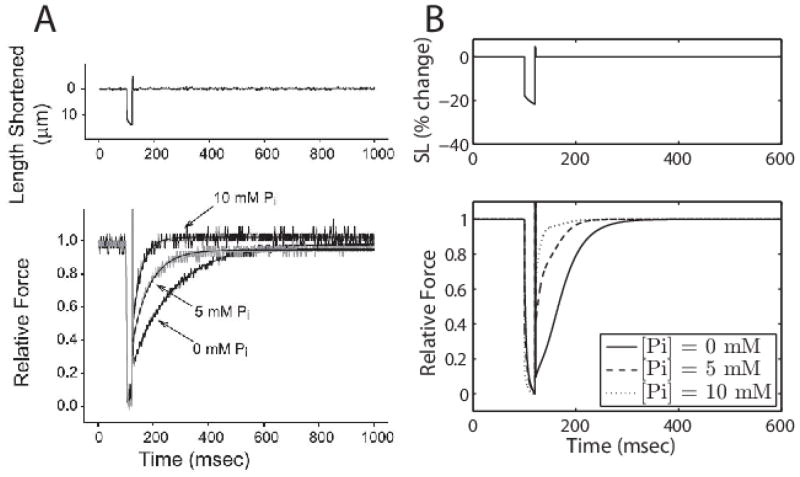
Despite extensive study over the past six decades the coupling of chemical reaction and mechanical processes in muscle dynamics is not well understood. We lack a theoretical description of how chemical processes (metabolite binding, ATP hydrolysis) influence and are influenced by mechanical processes (deformation and force generation). To address this need, a mathematical model of the muscle cross-bridge (XB) cycle based on Huxley's sliding filament theory is developed that explicitly accounts for the chemical transformation events and the influence of strain on state transitions. The model is identified based on elastic and viscous moduli data from mouse and rat myocardial strips over a range of perturbation frequencies, and MgATP and inorganic phosphate (Pi) concentrations. Simulations of the identified model reproduce the observed effects of MgATP and MgADP on the rate of force development. Furthermore, simulations reveal that the rate of force re-development measured in slack-restretch experiments is not directly proportional to the rate of XB cycling. For these experiments, the model predicts that the observed increase in the rate of force generation with increased Pi concentration is due to inhibition of cycle turnover by Pi. Finally, the model captures the observed phenomena of force yielding suggesting that it is a result of rapid detachment of stretched attached myosin heads.
Keywords: Cardiac muscle; Cross-bridge cycle; Force generation; Metabolites; Sinusoidal perturbation analysis; Viscoelasticity.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures











References
-
- Szent-Gyorgyi AG. The role of actin-myosin interaction in contraction. Symposia of the Society for Experimental Biology. 1968;22:17–42. - PubMed
-
- Huxley AF. Muscle structure and theories of contraction. Progress in biophysics and biophysical chemistry. 1957;7:255–318. - PubMed
-
- Huxley AF, Simmons RM. Proposed mechanism of force generation in striated muscle. Nature. 1971;233:533–8. - PubMed
-
- Cooke R. The mechanism of muscle contraction. CRC critical reviews in biochemistry. 1986;21:53–118. - PubMed
-
- Cooke R. Actomyosin interaction in striated muscle. Physiological reviews. 1997;77:671–97. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
